 Un Corazón está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre hacia los órganos periféricos. Cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente. Los ventrículos después aportan la principal fuerza del bombeo que impulsa la sangre:
Un Corazón está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre hacia los órganos periféricos. Cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente. Los ventrículos después aportan la principal fuerza del bombeo que impulsa la sangre:
1) hacia la circulación pulmonar por el ventrículo derecho.
2) hacia la circulación periférica por el ventrículo izquierdo.
Fisiología del músculo cardíaco
El corazón está formado por tres tiposde músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción.
 El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto que la duración de la contracción es mucho mayor. Las fibras especializadas de excitación y de conducción se contraen débilmente porque contienen pocas fibrillas contráctiles, pero presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción por todo el corazón, formando así un sistema excitador que controla el latido rítmico cardíaco.
El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto que la duración de la contracción es mucho mayor. Las fibras especializadas de excitación y de conducción se contraen débilmente porque contienen pocas fibrillas contráctiles, pero presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción por todo el corazón, formando así un sistema excitador que controla el latido rítmico cardíaco.
Anatomía fisiológica del músculo cardíaco
 El músculo cardíaco es estriado y tiene las miofibrillas típicas que contienen filamentos de actina y de miosina casi idénticos que se encuentran en el músculo esquelético; estos filamentos están unos al lado de otros
El músculo cardíaco es estriado y tiene las miofibrillas típicas que contienen filamentos de actina y de miosina casi idénticos que se encuentran en el músculo esquelético; estos filamentos están unos al lado de otros
y se deslizan entre sí durante la contracción de la misma
manera que ocurre en el músculo esquelético.
Músculo cardíaco como sincitio:
 Las zonas oscuras que atraviesan las fibras musculares cardíacas se denominan discos intercalados y son membranas celulares que separan las células musculares cardíacas individuales entre sí. En cada uno de los discos intercala-dos las membranas celulares se fusionan entre sí de tal manera que forman uniones comunicantes permeables que permiten una rápida difusión. Y desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular. el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una de ellas se excita el potencial de acción se propaga a todas. El corazón realmente está formado por dos sincitios:
Las zonas oscuras que atraviesan las fibras musculares cardíacas se denominan discos intercalados y son membranas celulares que separan las células musculares cardíacas individuales entre sí. En cada uno de los discos intercala-dos las membranas celulares se fusionan entre sí de tal manera que forman uniones comunicantes permeables que permiten una rápida difusión. Y desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular. el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una de ellas se excita el potencial de acción se propaga a todas. El corazón realmente está formado por dos sincitios:- El sincitio auricular, que forma las paredes de las dos aurículas.
- el sincitio ventricular, que forma las paredes de los dos ventrículos
Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV) entre las aurículas y los ventrículos.
Potenciales de acción en el músculo cardíaco
 El potencial de acción que se registra en una fibra muscular ventricular, es en promedio de aproximadamente 105 mV. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta. La presencia de esta meseta del potencial de acción hace que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético.
El potencial de acción que se registra en una fibra muscular ventricular, es en promedio de aproximadamente 105 mV. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta. La presencia de esta meseta del potencial de acción hace que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético.

En el músculo cardíaco, el potencial de acción está producido por la apertura de dos tipos de canales: 1) los canales rápidos de sodio.
2) canales lentos de calcio, que también se denominan canales de calcio-sodio.
Éstos últimos se abren con mayor lentitud y permanecen abiertos durante varias décimas de segundo, y esto mantiene un período prolongado de despolarización, dando lugar a la meseta del potencial de acción. Además, los iones calcio que entran durante esta fase de meseta activan el proceso contráctil del músculo.
La permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye; ésto se debe al exceso de flujo de entrada de calcio a través de los canales de calcio. La disminución de la permeabilidad al potasio reduce mucho el flujo de salida de iones potasio de carga positiva durante la meseta del potencial de acción y, por tanto, impide el regreso rápido del voltaje del potencial de acción a su nivel de reposo.
Velocidad de la conducción de las señales en el músculo cardíaco:
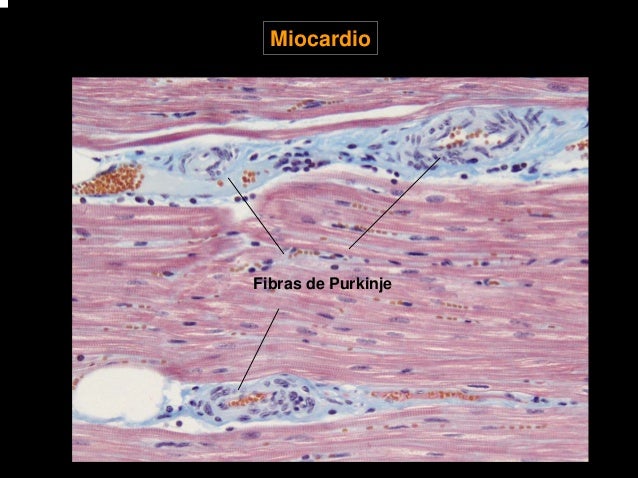
 La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de aproximadamente 0,3 a 0,5 m/s. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s.
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de aproximadamente 0,3 a 0,5 m/s. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s.
Período refractario del músculo cardíaco:
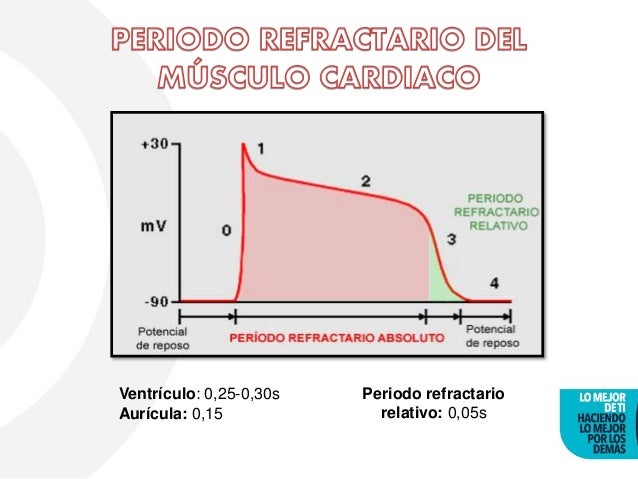 Es el intervalo de tiempo, durante el cual un impulso cardíaco normal no puede reexcitar una zona ya excitada de músculo cardíaco. El período refractario normal del ventrículo es de 0,25 a 0,30 s. Hay un período refractario relativo adicional de aproximadamente 0,05 s, durante el cual es más difícil de lo
Es el intervalo de tiempo, durante el cual un impulso cardíaco normal no puede reexcitar una zona ya excitada de músculo cardíaco. El período refractario normal del ventrículo es de 0,25 a 0,30 s. Hay un período refractario relativo adicional de aproximadamente 0,05 s, durante el cual es más difícil de lo
normal excitar el músculo pero, sin embargo, se puede excitar con una señal excitadora muy intensa. El período refractario del músculo auricular es mucho más corto que el de los ventrículos (0,15 s para las aurículas, y 0,25 a 0,30 s para los ventrículos).
Acoplamiento excitación-contracción: función de los iones calcio y de los túbulos transversos
El término acoplamiento excitación-contracción se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas del músculo se contraigan. Cuando un potencial de acción pasa sobre la membrana del músculo cardíaco el potencial de acción se propaga hacia la fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos (T); estos actúan sobre las membranas de los tubulos sarcoplásmicos longitudinales para producir la liberación de calcio. El calcio que entra en la célula activa después los canales de liberación de calcio, también denominados canales de receptor de rianodina, en la membrana del retículo sarcoplásmico, para activar la liberación de calcio en el sarcoplasma. Los iones calcio en el sarcoplasma interaccionan después con la troponina para iniciar la formación y contracción de puente transversal.
La cantidad de iones calcio en el sistema de los túbulos T depende en gran medida de la concentración de iones calcio en el líquido extracelular.
Al final de la meseta del potencial de acción cardíaco se interrumpe súbitamente el flujo de entrada de iones calcio hacia el interior de la fibra muscular. El transporte de calcio de nuevo al retículo sarcoplásmico se consigue con la ayuda de una bomba de calcio ATPasa.
El ciclo cardíaco
 Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco. Cada ciclo es iniciado por la generación espontánea de un potencial de acción en el nódulo sinusal; este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Debido a la posición de las estructuras, hay un retraso de más de 0,1 s durante el paso del impulso cardíaco desde las aurículas a los ventrículos lo que permite que las aurículas se contraigan antes de la contracción ventricular, bombeando de esta manera sangre hacia los ventrículos antes de que comience la intensa contracción ventricular.
Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco. Cada ciclo es iniciado por la generación espontánea de un potencial de acción en el nódulo sinusal; este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Debido a la posición de las estructuras, hay un retraso de más de 0,1 s durante el paso del impulso cardíaco desde las aurículas a los ventrículos lo que permite que las aurículas se contraigan antes de la contracción ventricular, bombeando de esta manera sangre hacia los ventrículos antes de que comience la intensa contracción ventricular.
Diástole y sístole
El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole. La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca. Para una frecuencia cardíaca normal de 72 latidos por minuto, la sístole comprende aproximadamente 0,4 del ciclo cardíaco completo.
Relación del electrocardiograma con el ciclo cardíaco:

El electrocardiograma es un conjunto de los voltajes eléctricos que genera el corazón, y son registrados mediante el electrocardiógrafo desde la superficie del cuerpo.
Función de las aurículas como bombas de cebado:
 La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. las aurículas actúan simplemente como bombas de cebado que aumentan la eficacia del bombeo ventricular hasta un 20%.
La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. las aurículas actúan simplemente como bombas de cebado que aumentan la eficacia del bombeo ventricular hasta un 20%.
Función de los ventrículos como bombas
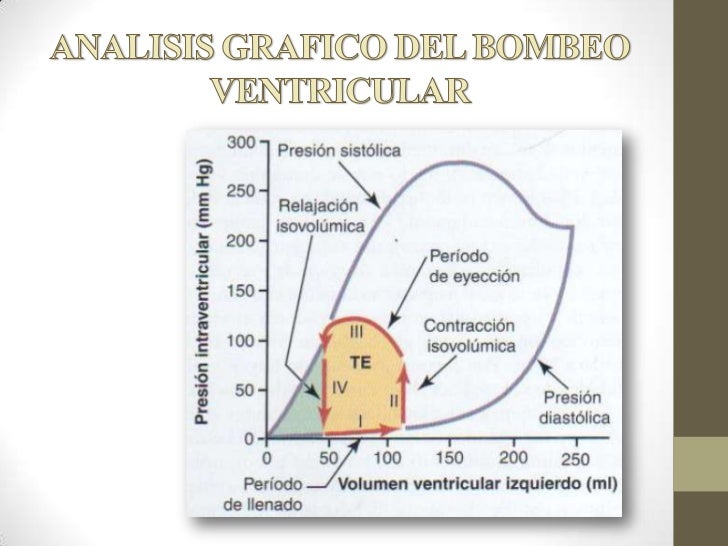
Llenado de los ventrículos durante la diástole: Durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas derecha e izquierda porque las válvulas AV están cerradas. Una vez que finaliza la sístole y las presiones ventriculares disminuyen de nuevo a sus valores diastólicos bajos, el aumento de presión que se ha generado en las aurículas durante la sístole ventricular inmediatamente abre las válvulas AV y permite que la sangre fluya rápidamente hacia los ventrículos, a este proceso se le llama período de llenado rápido de los ventrículos y dura aprox. el tercio de la diástole.
Vaciado de los ventrículos durante la sístole:
Período de contracción isovolumétrica (isométrica): Después del comienzo de la contracción ventricular se produce un aumento súbito de presión ventricular que hace que las válvulas se cierren. Después es necesario más tiempo para que se acumule más presió en el ventrículo, por lo que hay contracción pero no hay vaciado.

Período de eyección: Cuando la presión ventricular izquierda aumenta ligeramente por encima de 80 mmHg (y la presión ventricular derecha ligeramente por encima de 8 mmHg), las presiones ventriculares empiezan abrir las válvulas y se vacía la sangre en un 70% que corresponde al primer tercio del periodo de eyección por lo que se llama periodo de eyección rápida, y los dos finales se llaman periodo de eyección lenta que correponden al 30% de la sangre vaciada.

Período de relajación isovolumétrica (isométrica): Al final de la sístole comienza súbitamente la relajación ventricular, lo que permite que las presiones intraventriculares derecha e izquierda disminuyan rápidamente, por lo que el músculo cardiaco se relaja dando lugar a este período.
Volumen telediastólico: Durante la diástole, el llenado normal de los ventrículos aumenta el volumen de cada uno de los ventrículos hasta aproximadamente 110 a 120 ml.
Volumen sistólico: a medida que los ventrículos se vacían durante la sístole, el volumen disminuye aproximadamente 70 ml.
Volumen telesistólico: El volumen restante que queda en cada uno de los ventrículos, aproximadamente 40 a 50 ml.
Función de las válvulas
 Válvulas auriculoventriculares: Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole.
Válvulas auriculoventriculares: Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole.

 Función de los músculos papilares: se unen a los velos de las válvulas AV mediante las cuerdas tendinosas, se contraen cuando se contraen las paredes ventriculares, no contribuyen al cierre de las válvulas, por lo que tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular.
Función de los músculos papilares: se unen a los velos de las válvulas AV mediante las cuerdas tendinosas, se contraen cuando se contraen las paredes ventriculares, no contribuyen al cierre de las válvulas, por lo que tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular.
Válvula aórtica y de la arteria pulmonar:
https://blausen.com/es/video/como-funcionan-las-valvula-cardiacas/
Curva de presión aórtica:
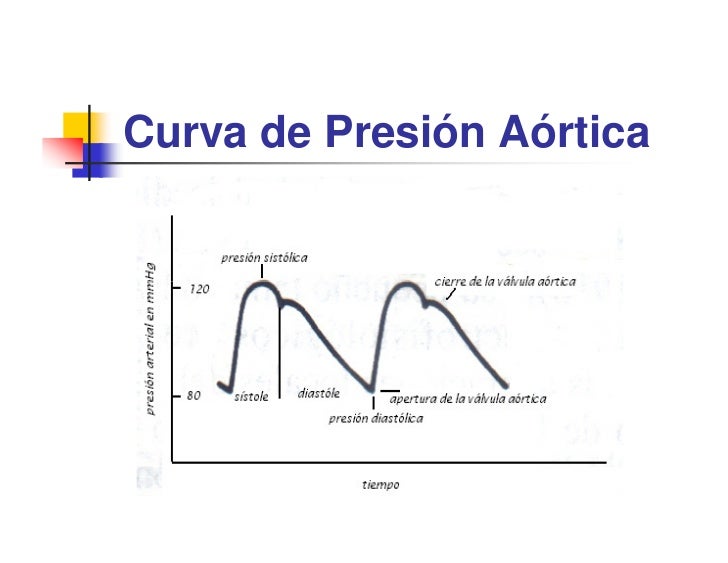 Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica. La entrada de sangre en las arterias hace que las paredes de las mismas se distiendan y que la presión aumente hasta aproximadamente 120 mmHg. Se produce la denominada incisura en la curva de presión aórtica cuando se cierra la válvula aórtica; está producida por un corto período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula, seguido por la interrupción súbita del flujo retrógrado.
Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica. La entrada de sangre en las arterias hace que las paredes de las mismas se distiendan y que la presión aumente hasta aproximadamente 120 mmHg. Se produce la denominada incisura en la curva de presión aórtica cuando se cierra la válvula aórtica; está producida por un corto período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula, seguido por la interrupción súbita del flujo retrógrado.
Relación de los tonos cardíacos con el bombeo cardíaco:

Generación de trabajo del corazón
El trabajo sistólico del corazón es la cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia las arterias. El trabajo minuto es la cantidad total de energía que se convierte en trabajo en 1 min; este parámetro es igual al trabajo sistólico multiplicado por la frecuencia cardíaca por minuto. El trabajo del corazón se usa para mover sangre desde las venas de baja presión a las arterias de alta presión. Esto se denomina trabajo volumen-presión o trabajo externo. También se utiliza para acelerar la sangre hasta su velocidad de eyección a través de las válvulas aórtica y pulmonar. Este es el componente de energía cinética del flujo sanguíneo del trabajo cardíaco.
El grado de tensión del músculo cuando comienza a contraerse se denomina precarga.
La carga contra la que el músculo ejerce su fuerza contráctil se denomina poscarga.
Energía química necesaria para la contracción cardíaca: la utilización de oxígeno por el corazón
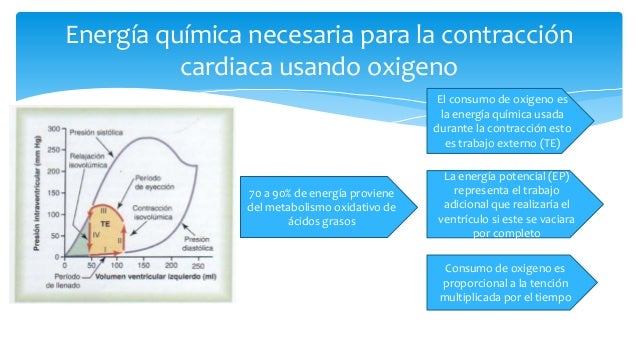
Regulación del bombeo cardiaco

Regulación intrínseca del bombeo cardíaco: el mecanismo de Frank-Starling
La cantidad de sangre que bombea el corazón cada minuto está determinada, por lo común, casi totalmente por la velocidad del flujo sanguíneo hacia el corazón desde las venas, que se denomina retorno venoso. Es decir, todos los tejidos periféricos del cuerpo controlan su propio flujo sanguíneo local, y todos los flujos tisulares locales se combinan y regresan a través de las venas hacia la aurícula derecha.
Curvas de función ventricular

Las curvas de función ventricular son otra forma de expresar el mecanismo de Frank-Starling del corazón. Es decir, a medida que los ventrículos se llenan en respuesta a unas presiones auriculares más altas, se produce aumento del volumen de los dos ventrículos y de la fuerza de la contracción del músculo cardíaco, lo que hace que el corazón bombee mayores cantidades de sangre hacia las arterias.
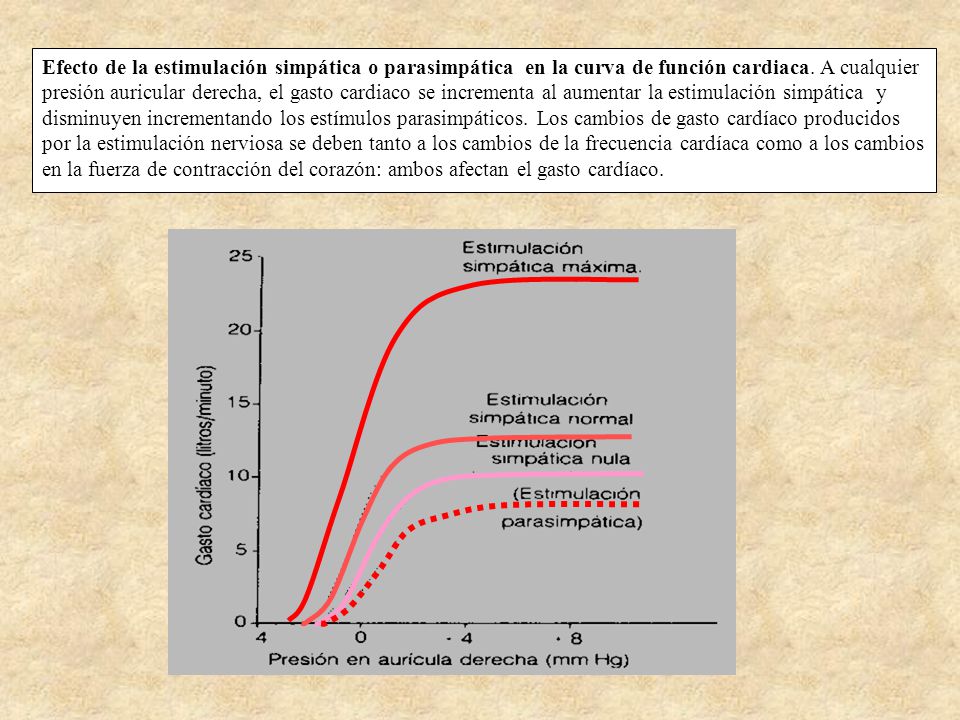
Control del corazón por los nervios simpáticos y parasimpáticos
 Mecanismos de excitación del corazón por los nervios simpáticos: La estimulación simpática aumenta la fuerza de la contracción cardíaca hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección; la inhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón, este fenómeno produce reducción tanto de la frecuencia cardíaca como de la fuerza de la contracción del músculo ventricular.
Mecanismos de excitación del corazón por los nervios simpáticos: La estimulación simpática aumenta la fuerza de la contracción cardíaca hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección; la inhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón, este fenómeno produce reducción tanto de la frecuencia cardíaca como de la fuerza de la contracción del músculo ventricular.
Estimulación parasimpática (vagal) del corazón: La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos. Las fibras vagales se distribuyen principalmente por las aurículas y no mucho en los ventrículos.
Efecto de los iones potasio y calcio sobre la función cardíaca
- El exceso de potasio hace que el corazón esté dilatado y flácido, y también reduce la frecuencia cardíaca. Grandes cantidades también pueden bloquear la conducción del impulso cardíaco desde las aurículas hacia los ventrículos a través del haz AV.
- El exceso de iones calcio hace que el corazón progrese hacia una contracción espástica, por el contrario, el déficit de iones calcio produce flacidez cardíaca.
Efecto de la temperatura sobre la función cardíaca
 El aumento de la temperatura corporal, produce un gran aumento de la frecuencia cardíaca, a veces hasta del doble del valor normal. La disminución de la temperatura produce una gran disminución de la frecuencia cardíaca, que puede disminuir hasta sólo algunos latidos por minuto.
El aumento de la temperatura corporal, produce un gran aumento de la frecuencia cardíaca, a veces hasta del doble del valor normal. La disminución de la temperatura produce una gran disminución de la frecuencia cardíaca, que puede disminuir hasta sólo algunos latidos por minuto.

 Músculo liso multiunitario: Este tipo de músculo liso está formado por fibras musculares lisas separadas y discretas. Cada fibra actúa independientemente y está inervada por una sóla terminación nerviosa. Su superficie externa está cubierta por una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí. La característica más importante de las fibras musculares lisas multiunitarias es que cada una de las fibras se puede contraer independientemente de las demás, y su control se ejerce principalmente por señales nerviosas.
Músculo liso multiunitario: Este tipo de músculo liso está formado por fibras musculares lisas separadas y discretas. Cada fibra actúa independientemente y está inervada por una sóla terminación nerviosa. Su superficie externa está cubierta por una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí. La característica más importante de las fibras musculares lisas multiunitarias es que cada una de las fibras se puede contraer independientemente de las demás, y su control se ejerce principalmente por señales nerviosas.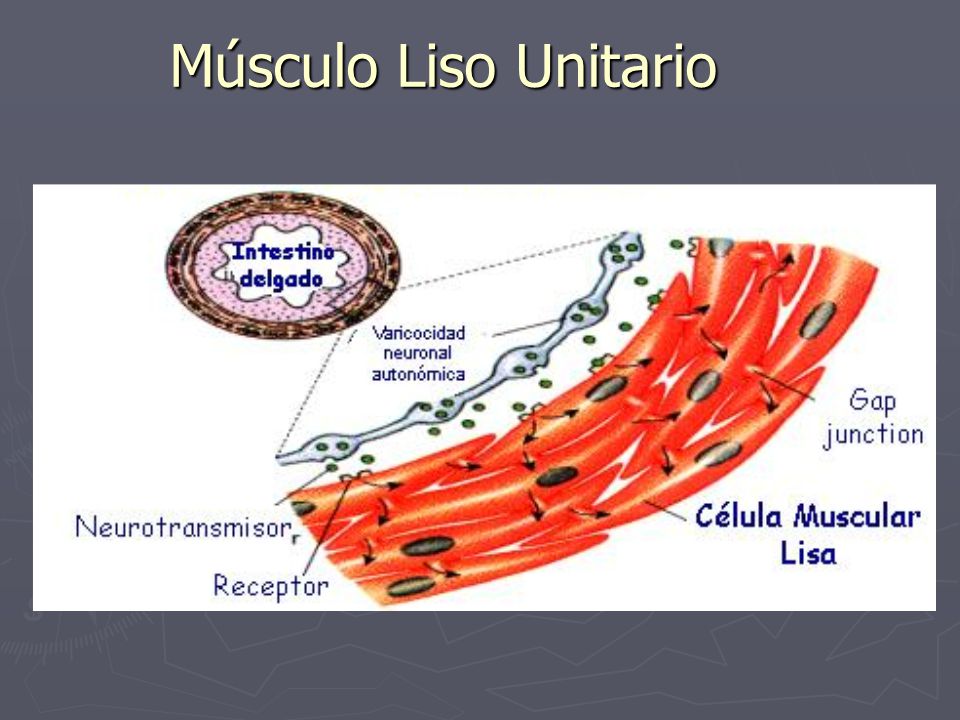 Músculo liso unitario (músculo liso sincitial o músculo liso visceral) : Es una masa de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad. Las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra, de modo que los potenciales de acción o el flujo iónico simple sin potenciales de acción puede viajar desde una fibra a otra y hacer que las fibras musculares se contraigan simultáneamente.
Músculo liso unitario (músculo liso sincitial o músculo liso visceral) : Es una masa de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad. Las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra, de modo que los potenciales de acción o el flujo iónico simple sin potenciales de acción puede viajar desde una fibra a otra y hacer que las fibras musculares se contraigan simultáneamente.
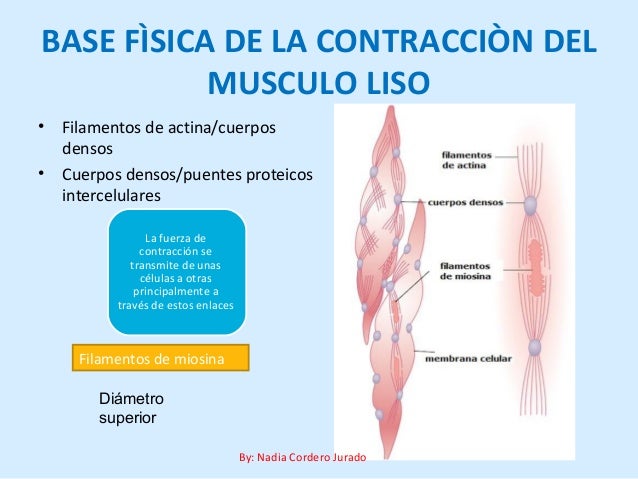

 Una caracteristica importante del músculo unitario visceral es su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado. Existen unos fenómenos que se denominan tensión-relajación y tensión-relajación inversa.
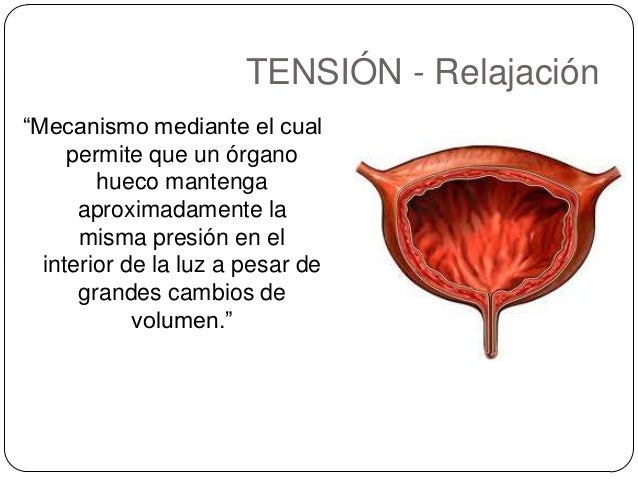
Una caracteristica importante del músculo unitario visceral es su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado. Existen unos fenómenos que se denominan tensión-relajación y tensión-relajación inversa. el estímulo que inicia la mayor parte de las contracciones del músculo liso es un aumento de los iones calcio en el medio intracelular.
el estímulo que inicia la mayor parte de las contracciones del músculo liso es un aumento de los iones calcio en el medio intracelular. La miosina fosfatasa, que está localizada en el citosol de la célula muscular lisa y que escinde el fosfato de la cadena ligera reguladora. Después se interrumpe el ciclo y finaliza la contracción. Por tanto, el tiempo necesario para la relajación de
La miosina fosfatasa, que está localizada en el citosol de la célula muscular lisa y que escinde el fosfato de la cadena ligera reguladora. Después se interrumpe el ciclo y finaliza la contracción. Por tanto, el tiempo necesario para la relajación de
 Las fibras nerviosas autónomas que inervan el músculo liso generalmente se ramifican de manera difusa encima de una lámina de fibras musculares. En la mayor parte de los casos estas fibras no hacen contacto directo con la membrana de las células de las fibras musculares lisas, sino que forman las denominadas uniones difusas que secretan su sustancia transmisora hacia el recubrimiento de matriz del músculo liso cuando hay muchas capas de células musculares, las fibras nerviosas con frecuencia inervan sólo la capa externa.
Las fibras nerviosas autónomas que inervan el músculo liso generalmente se ramifican de manera difusa encima de una lámina de fibras musculares. En la mayor parte de los casos estas fibras no hacen contacto directo con la membrana de las células de las fibras musculares lisas, sino que forman las denominadas uniones difusas que secretan su sustancia transmisora hacia el recubrimiento de matriz del músculo liso cuando hay muchas capas de células musculares, las fibras nerviosas con frecuencia inervan sólo la capa externa.
 Despolarización del músculo liso multiunitario sin potenciales de acción: Las fibras musculares lisas del músculo liso multiunitario como el músculo del iris del ojo o el músculo erector de cada uno de los cabellos se contraen sobre todo en respuesta a estímulos nerviosos. Las terminaciones nerviosas secretan acetilcolina en algunos y noradrenalina en el caso de otros. En ambos casos, las sustancias transmisoras producen despolarización de la membrana del músculo liso, y esto a su vez produce la contracción.
Despolarización del músculo liso multiunitario sin potenciales de acción: Las fibras musculares lisas del músculo liso multiunitario como el músculo del iris del ojo o el músculo erector de cada uno de los cabellos se contraen sobre todo en respuesta a estímulos nerviosos. Las terminaciones nerviosas secretan acetilcolina en algunos y noradrenalina en el caso de otros. En ambos casos, las sustancias transmisoras producen despolarización de la membrana del músculo liso, y esto a su vez produce la contracción.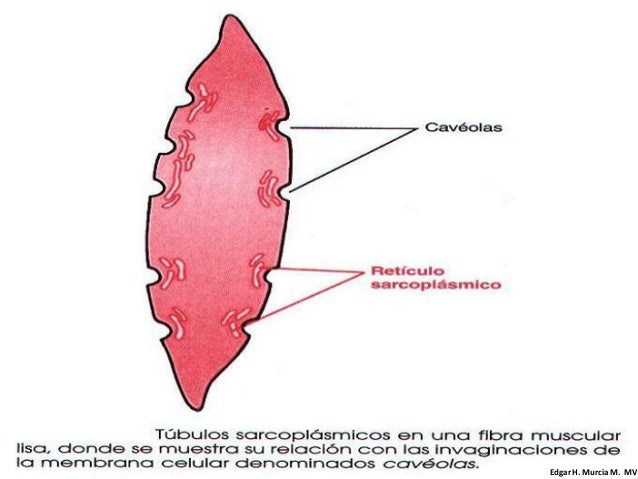 El retículo sarcoplásmico, que aporta prácticamente los iones calcio para la contracción del músculo esquelético, está poco desarrollado en la mayor parte del músculo liso. El tiempo necesario para que se produzca esta difusión es en promedio de entre 200 y 300 ms y se denomina período de latencia antes de que comience la contracción. Este período de latencia es aproximadamente 50 veces mayor para la contracción del músculo liso que para la del músculo esquelético.
El retículo sarcoplásmico, que aporta prácticamente los iones calcio para la contracción del músculo esquelético, está poco desarrollado en la mayor parte del músculo liso. El tiempo necesario para que se produzca esta difusión es en promedio de entre 200 y 300 ms y se denomina período de latencia antes de que comience la contracción. Este período de latencia es aproximadamente 50 veces mayor para la contracción del músculo liso que para la del músculo esquelético.
 La fibra nerviosa forma un complejo de terminaciones nerviosas ramificadas que se invaginan en la superficie de la fibra muscular, pero que permanecen fuera de la membrana plasmática de la misma. Toda la estructura se denomina placa motora terminal. Esta cubierta por una o más células de Schwann que la aíslan de los líquidos circundantes.
La fibra nerviosa forma un complejo de terminaciones nerviosas ramificadas que se invaginan en la superficie de la fibra muscular, pero que permanecen fuera de la membrana plasmática de la misma. Toda la estructura se denomina placa motora terminal. Esta cubierta por una o más células de Schwann que la aíslan de los líquidos circundantes. La membrana invaginada se denomina gotiera sináptica o valle sináptico.
La membrana invaginada se denomina gotiera sináptica o valle sináptico. La acetilcolina excita a la membrana de la fibra muscular. La acetilcolina se sintetiza en el citoplasma de la terminación, se absorbe rápidamente hacia el interior de muchas pequeñas vesículas sinápticas, de las que normalmente hay aproxi-madamente 300.000 en las terminaciones de una única placa terminal. En el espacio sináptico hay grandes cantidades de la enzima acetilcolinesterasa, que destruye la acetilcolina después de que la hayan liberado las vesículas sinápticas.
La acetilcolina excita a la membrana de la fibra muscular. La acetilcolina se sintetiza en el citoplasma de la terminación, se absorbe rápidamente hacia el interior de muchas pequeñas vesículas sinápticas, de las que normalmente hay aproxi-madamente 300.000 en las terminaciones de una única placa terminal. En el espacio sináptico hay grandes cantidades de la enzima acetilcolinesterasa, que destruye la acetilcolina después de que la hayan liberado las vesículas sinápticas. En la superficie interna de la membrana neural hay barras densas lineales, a los dos lados de estas hay canales de calcio activados por voltaje; cuando un potencial de acción se propaga por la terminación, estos canales se abren y permiten que iones calcio difundan desde el espacio sináptico hacia el interior de la terminación nerviosa, teoricamente los iones calcio ejercen una influencia de atracción sobre las vesículas de acetilcolina, desplazándolas hacia la membrana neural; así las vesículas se fusionan con la membrana neural y vacían su acetilcolina hacia el espacio sináptico mediante el proceso de exocitosis.
En la superficie interna de la membrana neural hay barras densas lineales, a los dos lados de estas hay canales de calcio activados por voltaje; cuando un potencial de acción se propaga por la terminación, estos canales se abren y permiten que iones calcio difundan desde el espacio sináptico hacia el interior de la terminación nerviosa, teoricamente los iones calcio ejercen una influencia de atracción sobre las vesículas de acetilcolina, desplazándolas hacia la membrana neural; así las vesículas se fusionan con la membrana neural y vacían su acetilcolina hacia el espacio sináptico mediante el proceso de exocitosis. proteico formado por cinco subunidades proteicas, dos proteínas alfa y una proteína beta, una delta y una gamma. Estas moléculas proteicas atraviesan la membrana, y están dispuestas en círculo para formar un canal tubularque permanece cerrado hasta que dos moléculas de acetilcolina se unen a dos proteínas alfa.
proteico formado por cinco subunidades proteicas, dos proteínas alfa y una proteína beta, una delta y una gamma. Estas moléculas proteicas atraviesan la membrana, y están dispuestas en círculo para formar un canal tubularque permanece cerrado hasta que dos moléculas de acetilcolina se unen a dos proteínas alfa. El canal activado por acetilcolina tiene un diámetro de aproximadamente 0,65 nm, que es lo suficientemente grande como para permitir que los iones positivos importantes Na, K y Ca se muevan con facilidad a través de la abertura. los iones cloruro, no lo atraviesan debido a las intensas cargas negativas de la abertura del canal que las repelen; el principal efecto de la apertura de los canales activados por la acetilcolina es permitir que grandes cantidades de iones sodio entren al interior de la fibra, desplazando con ellos grandes números de cargas positivas. Esto genera un cambio de potencial positivo local en la membrana de la fibra muscular, denominado potencial de la placa terminal.
El canal activado por acetilcolina tiene un diámetro de aproximadamente 0,65 nm, que es lo suficientemente grande como para permitir que los iones positivos importantes Na, K y Ca se muevan con facilidad a través de la abertura. los iones cloruro, no lo atraviesan debido a las intensas cargas negativas de la abertura del canal que las repelen; el principal efecto de la apertura de los canales activados por la acetilcolina es permitir que grandes cantidades de iones sodio entren al interior de la fibra, desplazando con ellos grandes números de cargas positivas. Esto genera un cambio de potencial positivo local en la membrana de la fibra muscular, denominado potencial de la placa terminal.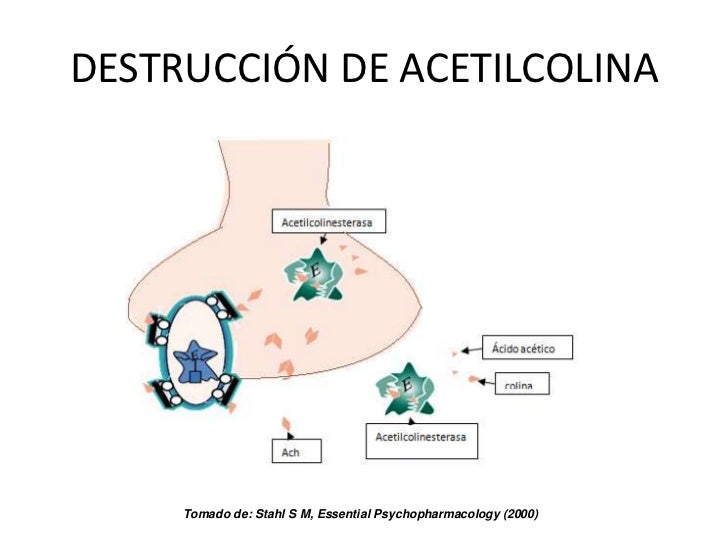
 La rápida entrada de iones sodio en la fibra muscular cuando se abren los canales activados por acetilcolina hace que el potencial eléctrico en el interior de la fibra en la zona local de la placa terminal aumente en dirección positiva hasta 50 a 75 mV, generando un potencial local denominado potencial de la placa terminal.
La rápida entrada de iones sodio en la fibra muscular cuando se abren los canales activados por acetilcolina hace que el potencial eléctrico en el interior de la fibra en la zona local de la placa terminal aumente en dirección positiva hasta 50 a 75 mV, generando un potencial local denominado potencial de la placa terminal. Curare es un fármaco que bloquea la acción activadora de la acetilcolina sobre los canales de acetilcolina compitiendo con los puntos del receptor de acetilcolina.
Curare es un fármaco que bloquea la acción activadora de la acetilcolina sobre los canales de acetilcolina compitiendo con los puntos del receptor de acetilcolina. Toxina botulínica es un veneno bacteriano que reduce la magnitud de la liberación de acetilcolina por las terminaciones nerviosas.
Toxina botulínica es un veneno bacteriano que reduce la magnitud de la liberación de acetilcolina por las terminaciones nerviosas. Cada impulso que llega a la unión neuromuscular produce un potencial de la placa terminal mayor que el necesario para estimular la fibra nerviosa. Por tanto, se dice que la unión neuromuscular normal tiene un elevado factor de seguridad. La estimulación de la fibra nerviosa a frecuencias mayores de 100 veces por segundo durante varios minutos disminuye tanto el número de vesículas de acetilcolina que los impulsos no pueden pasar hacia la fibra nerviosa. Esto se denomina fatiga de la unión neuromuscular y es el mismo efecto que produce fatiga de las sinapsis en el sistema nervioso central cuando las sinapsis son sobreexcitadas.
Cada impulso que llega a la unión neuromuscular produce un potencial de la placa terminal mayor que el necesario para estimular la fibra nerviosa. Por tanto, se dice que la unión neuromuscular normal tiene un elevado factor de seguridad. La estimulación de la fibra nerviosa a frecuencias mayores de 100 veces por segundo durante varios minutos disminuye tanto el número de vesículas de acetilcolina que los impulsos no pueden pasar hacia la fibra nerviosa. Esto se denomina fatiga de la unión neuromuscular y es el mismo efecto que produce fatiga de las sinapsis en el sistema nervioso central cuando las sinapsis son sobreexcitadas. 1. Se forman pequeñas vesículas en el aparato de Golgi, las cuales son transportadas hasta la unión neuromuscular en las terminaciones nerviosas periféricas; se acumulan aprox. 300,000.
1. Se forman pequeñas vesículas en el aparato de Golgi, las cuales son transportadas hasta la unión neuromuscular en las terminaciones nerviosas periféricas; se acumulan aprox. 300,000.
 3. Cuando un potencial de acción llega a la terminación nerviosa, abre canales de calcio en la membrana de la terminación nerviosa, y aumenta la velocidad de fusión de las vesículas de acetilcolina, lo que hace que muchas de las vesículas se rompan, permitiendo la exocitosis de la acetilcolina hacia el espacio sináptico, después la acetilcolina es dividida por la acetilcolinesterasa en ion de acetato y colina, y la colina se reabsorbe activamente en la terminación neural para ser reutilizada para formar de nuevo acetilcolina.
3. Cuando un potencial de acción llega a la terminación nerviosa, abre canales de calcio en la membrana de la terminación nerviosa, y aumenta la velocidad de fusión de las vesículas de acetilcolina, lo que hace que muchas de las vesículas se rompan, permitiendo la exocitosis de la acetilcolina hacia el espacio sináptico, después la acetilcolina es dividida por la acetilcolinesterasa en ion de acetato y colina, y la colina se reabsorbe activamente en la terminación neural para ser reutilizada para formar de nuevo acetilcolina. 4. Al terminar los potenciales de acción aparecen hendiduras revestidas en la membrana de la terminación nerviosa, producidas por las proteínas, especialmente la proteína clatrina; las proteínas se contraen y hacen que las hendiduras se rompan hacia el interior de la membrana, formando de esta manera nuevas vesículas, y se repite el proceso.
4. Al terminar los potenciales de acción aparecen hendiduras revestidas en la membrana de la terminación nerviosa, producidas por las proteínas, especialmente la proteína clatrina; las proteínas se contraen y hacen que las hendiduras se rompan hacia el interior de la membrana, formando de esta manera nuevas vesículas, y se repite el proceso.
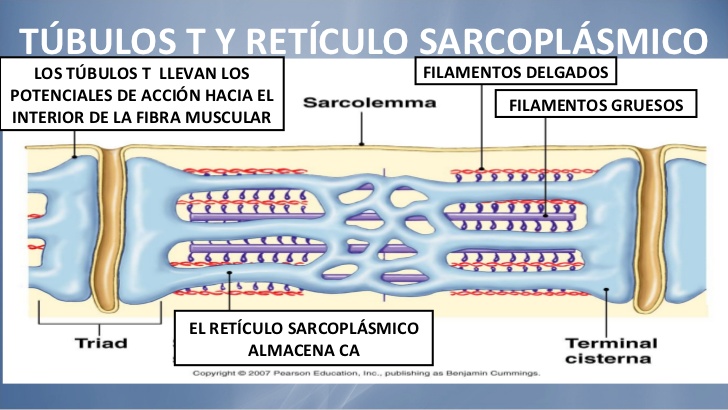 Los túbulos T son realmente extensiones internas de la membrana celular; por tanto, cuando un potencial de acción se propaga por la membrana de una fibra muscular, también se propaga un cambio de potencial a lo largo de los túbulos T hacia las zonas profundas del interior de la fibra muscular. De esta manera las corrientes eléctricas que rodean a estos túbulos T producen la contracción muscular.
Los túbulos T son realmente extensiones internas de la membrana celular; por tanto, cuando un potencial de acción se propaga por la membrana de una fibra muscular, también se propaga un cambio de potencial a lo largo de los túbulos T hacia las zonas profundas del interior de la fibra muscular. De esta manera las corrientes eléctricas que rodean a estos túbulos T producen la contracción muscular.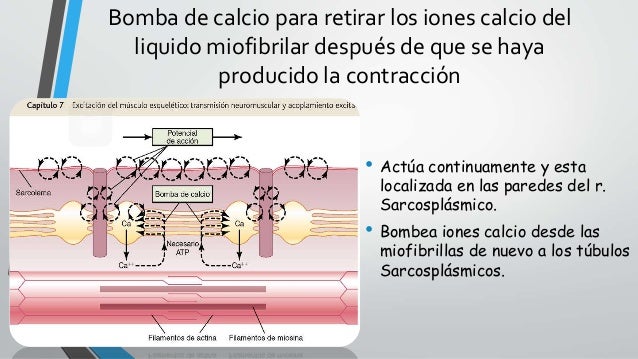 Esta bomba puede concentrar los iones calcio aproximada-mente 10.000 veces en el
Esta bomba puede concentrar los iones calcio aproximada-mente 10.000 veces en el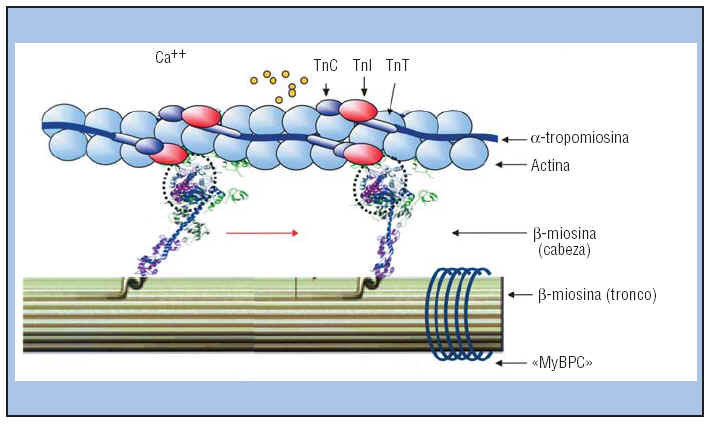 La concentración normal en estado de reposo (<10–7 molar) de los iones calcio en el citosol que baña las miofibrillas es demasiado pequeña como para producir una contracción. El complejo troponina-tropomiosina mantiene inhibidos los filamentos de actina y mantiene el estado relajado del músculo. Por el contrario, la excitación completa del sistema del túbulo T y del retículo sarcoplásmico da lugar a una liberación de iones calcio suficiente como para aumentar la concentración en el líquido miofibrilar; a esto se le llama pulso de calcio en la fibra muscular esquelética. Durante este pulso de calcio se produce la contracción muscular.
La concentración normal en estado de reposo (<10–7 molar) de los iones calcio en el citosol que baña las miofibrillas es demasiado pequeña como para producir una contracción. El complejo troponina-tropomiosina mantiene inhibidos los filamentos de actina y mantiene el estado relajado del músculo. Por el contrario, la excitación completa del sistema del túbulo T y del retículo sarcoplásmico da lugar a una liberación de iones calcio suficiente como para aumentar la concentración en el líquido miofibrilar; a esto se le llama pulso de calcio en la fibra muscular esquelética. Durante este pulso de calcio se produce la contracción muscular.


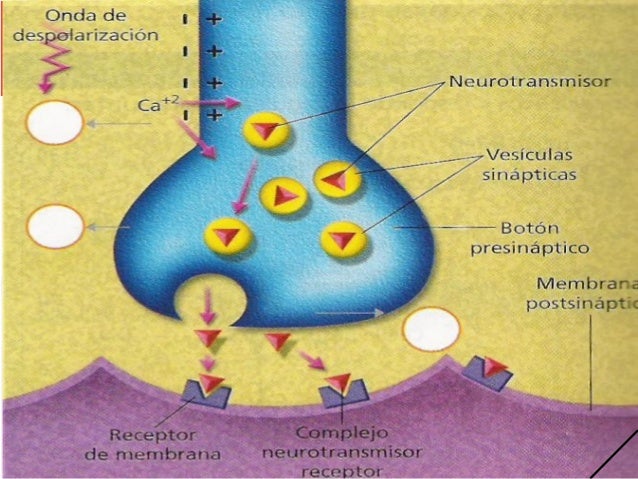 Una sinapsis es posible gracias al botón sinaptico, el cual es un conjunto de ramificaciones al final del axón que permiten que el impulso nervioso se propague, también conocido como botón terminal, ya que se encuentra en el extremo de una rama terminal del axón de una neurona presináptica. En el botón existen vesículas sinápticas que contienen los neurotransmisores que los libera para producir impulsos hacia otra neurona, músculo, etc.
Una sinapsis es posible gracias al botón sinaptico, el cual es un conjunto de ramificaciones al final del axón que permiten que el impulso nervioso se propague, también conocido como botón terminal, ya que se encuentra en el extremo de una rama terminal del axón de una neurona presináptica. En el botón existen vesículas sinápticas que contienen los neurotransmisores que los libera para producir impulsos hacia otra neurona, músculo, etc. Placa neuromuscular (unión neuromuscular): es la sinapsis entre una neurona motora y un músculo, gracias a los impulsos transmitidos, el músculo puede contraerse o relajarse. Los botones terminales de las neuronas se conectan con las placas terminales motoras. Éstas últimas se refieren a la membrana que recibe los impulsos nerviosos de una placa neuromuscular.
Placa neuromuscular (unión neuromuscular): es la sinapsis entre una neurona motora y un músculo, gracias a los impulsos transmitidos, el músculo puede contraerse o relajarse. Los botones terminales de las neuronas se conectan con las placas terminales motoras. Éstas últimas se refieren a la membrana que recibe los impulsos nerviosos de una placa neuromuscular. – Una neurona motora (motoneurona). Esta neurona se denomina presináptica porque emite impulsos nerviosos o potenciales de acción. Específicamente, los impulsos nerviosos viajan por el axón de esta neurona hasta el botón terminal que se sitúa muy cerca del músculo.
– Una neurona motora (motoneurona). Esta neurona se denomina presináptica porque emite impulsos nerviosos o potenciales de acción. Específicamente, los impulsos nerviosos viajan por el axón de esta neurona hasta el botón terminal que se sitúa muy cerca del músculo.
 Las fibras musculares que se inervan en la placa neuromuscular se llaman fibras musculares extrafusales. Son las controladas por motoneuronas alfa y son responsables de la fuerza que surge de la contracción de un músculo esquelético.
Las fibras musculares que se inervan en la placa neuromuscular se llaman fibras musculares extrafusales. Son las controladas por motoneuronas alfa y son responsables de la fuerza que surge de la contracción de un músculo esquelético.

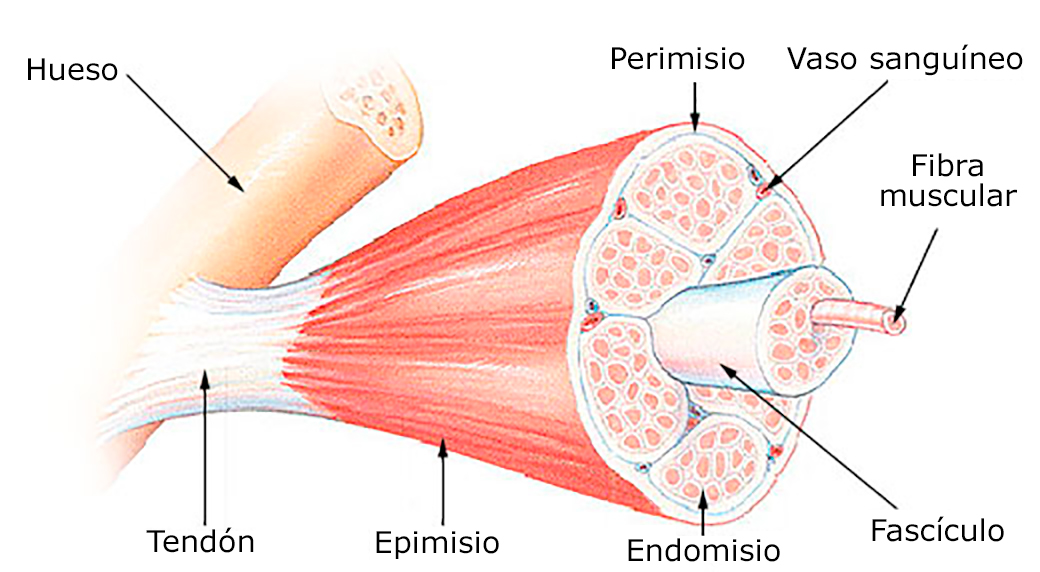 Los músculos esqueléticos están formados por fibras cuyo diámetro oscila entre los 10 y 80 micrómetros. En su mayoría, las fibras de los músculos esqueléticos se extienden a lo largo de toda la longitud del músculo. Todas las fibras habitualmente están inervadas sólo por una terminación nerviosa, que está localizada cerca del punto medio de la misma.
Los músculos esqueléticos están formados por fibras cuyo diámetro oscila entre los 10 y 80 micrómetros. En su mayoría, las fibras de los músculos esqueléticos se extienden a lo largo de toda la longitud del músculo. Todas las fibras habitualmente están inervadas sólo por una terminación nerviosa, que está localizada cerca del punto medio de la misma.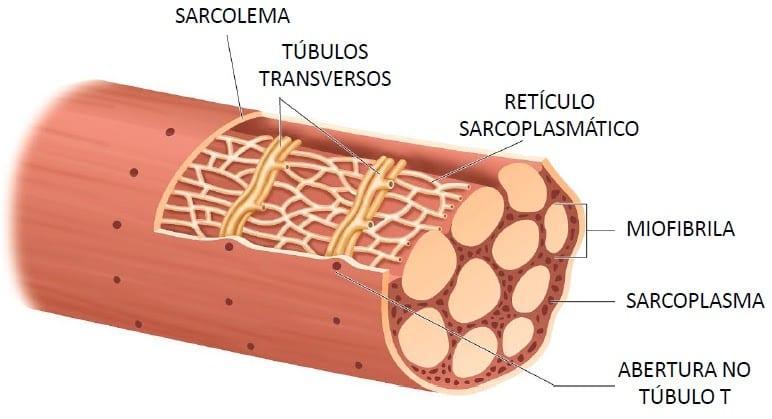 El sarcolema está formado por una membrana celular verdadera, denominada membrana plasmática, y una cubierta externa formada por una capa delgada de polisa-cárido que contiene numerosas fibrillas delgadas de colágeno. En ambos extremos de la fibra muscular, la capa superficial del sarcolema se fusiona con una fibra tendinosa. Las fibras tendinosas a su vez se agrupan en haces para formar los tendones musculares, que después se insertan en los huesos.
El sarcolema está formado por una membrana celular verdadera, denominada membrana plasmática, y una cubierta externa formada por una capa delgada de polisa-cárido que contiene numerosas fibrillas delgadas de colágeno. En ambos extremos de la fibra muscular, la capa superficial del sarcolema se fusiona con una fibra tendinosa. Las fibras tendinosas a su vez se agrupan en haces para formar los tendones musculares, que después se insertan en los huesos. Cada fibra muscular va a estar conformada por una serie de miofibrillas, cada una de ellas formada por filamentos de actina y de miosina (grandes moléculas proteicas encargadas de la contracción). los filamentos de miosina y de actina se unen o entrelazan parcialmente y de esta manera hacen que las miofibrillas tengan bandas claras y oscuras alternas. Las bandas claras contienen sólo filamentos de actina y se denominan bandas I. Las bandas oscuras contienen filamentos de miosina, y se denominan bandas A. Estas bandas alternadas le dan al músculo esquelético y cardiaco su aspecto estriado. Hay pequeñas proyecciones que se originan en los lados de los filamentos de miosina que se denominan puentes cruzados. La interacción entre estos puentes cruzados y los filamentos de actina produce la contracción.
Cada fibra muscular va a estar conformada por una serie de miofibrillas, cada una de ellas formada por filamentos de actina y de miosina (grandes moléculas proteicas encargadas de la contracción). los filamentos de miosina y de actina se unen o entrelazan parcialmente y de esta manera hacen que las miofibrillas tengan bandas claras y oscuras alternas. Las bandas claras contienen sólo filamentos de actina y se denominan bandas I. Las bandas oscuras contienen filamentos de miosina, y se denominan bandas A. Estas bandas alternadas le dan al músculo esquelético y cardiaco su aspecto estriado. Hay pequeñas proyecciones que se originan en los lados de los filamentos de miosina que se denominan puentes cruzados. La interacción entre estos puentes cruzados y los filamentos de actina produce la contracción. La porción de la miofibrilla que está entre dos discos Z se llama sarcómero, y cuando la fibra muscilar se contrae la longitud del sarcómero es de 2 micrómetros aprox. y es a esta longitud el músculo es capaz de generar su máxima fuerza de contracción.
La porción de la miofibrilla que está entre dos discos Z se llama sarcómero, y cuando la fibra muscilar se contrae la longitud del sarcómero es de 2 micrómetros aprox. y es a esta longitud el músculo es capaz de generar su máxima fuerza de contracción.
 En el estado relajado, los extremos de los filamentos de actina que se extienden entre dos discos Z sucesivos apenas comienzan a superponerse entre sí. Por el contrario, en el estado contraído estos filamentos de actina han sido traccionados hacia dentro entre los filamentos de miosina, de modo que sus extremos se superponen entre sí en su máxima extensión. Así, la contracción muscular se produce por un mecanismo de deslizamiento de los filamentos. Este fenómeno está producido por las fuerzas que se generan por la interacción de los puentes cruzados que van desde los filamentos de miosina a los filamentos de actina.
En el estado relajado, los extremos de los filamentos de actina que se extienden entre dos discos Z sucesivos apenas comienzan a superponerse entre sí. Por el contrario, en el estado contraído estos filamentos de actina han sido traccionados hacia dentro entre los filamentos de miosina, de modo que sus extremos se superponen entre sí en su máxima extensión. Así, la contracción muscular se produce por un mecanismo de deslizamiento de los filamentos. Este fenómeno está producido por las fuerzas que se generan por la interacción de los puentes cruzados que van desde los filamentos de miosina a los filamentos de actina.
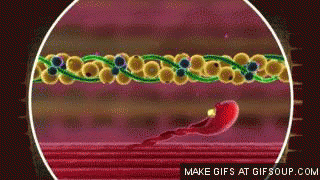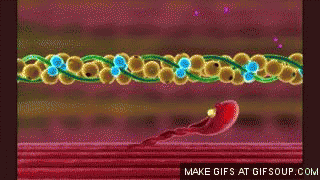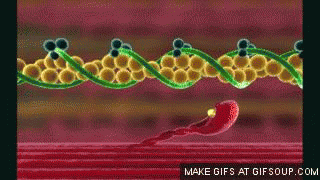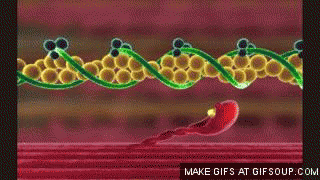 En estado de reposo las moléculas de tropomiosina recubren los puntos activos de las hebras de actina, de modo que no se puede producir atracción entre los filamentos de actina y de miosina para producir la contracción.
En estado de reposo las moléculas de tropomiosina recubren los puntos activos de las hebras de actina, de modo que no se puede producir atracción entre los filamentos de actina y de miosina para producir la contracción.
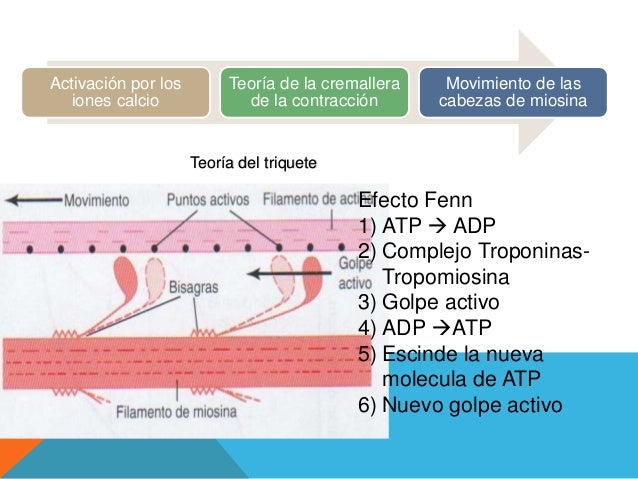 Tan pronto como el filamento de actina es activado por los iones calcio, las cabezas de los puentes cruzados de los filamentos de miosina son atraídos hacia los puntos activos del filamento de actina y de algún modo esto hace que se produzca la contracción.
Tan pronto como el filamento de actina es activado por los iones calcio, las cabezas de los puentes cruzados de los filamentos de miosina son atraídos hacia los puntos activos del filamento de actina y de algún modo esto hace que se produzca la contracción. La tensión desarrollada en un músculo en contra-cción va a estar determinada por la superposición de los filamentos de actina y miosina.
La tensión desarrollada en un músculo en contra-cción va a estar determinada por la superposición de los filamentos de actina y miosina.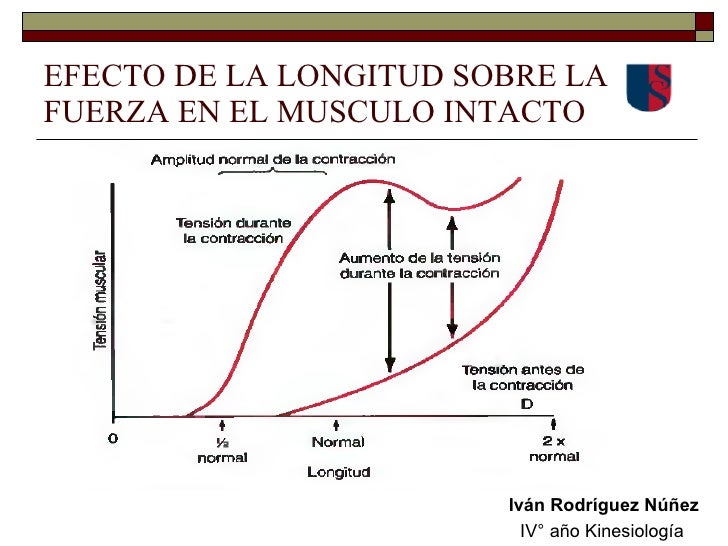 Cuando el músculo está en su longitud normal en reposo, que corresponde a una longitud del sarcómero de aproximadamente 2 micrómetros, se contrae con una fuerza de contracción próxima a la fuerza máxima cuando es activado. Sin embargo, el aumento de la tensión que se produce durante la contracción, denominado tensión activa, se reduce a medida que el músculo es distendido más allá de su longitud normal, es decir, hasta una longitud del sarcómero mayor de aproximadamente 2,2 micrómetros.
Cuando el músculo está en su longitud normal en reposo, que corresponde a una longitud del sarcómero de aproximadamente 2 micrómetros, se contrae con una fuerza de contracción próxima a la fuerza máxima cuando es activado. Sin embargo, el aumento de la tensión que se produce durante la contracción, denominado tensión activa, se reduce a medida que el músculo es distendido más allá de su longitud normal, es decir, hasta una longitud del sarcómero mayor de aproximadamente 2,2 micrómetros. Un músculo esquelético se contrae muy rápido cuando no tiene ninguna carga, y va disminuyendo su velocidad conforme la carga va a aumentando. cuando la carga ha aumentado hasta la fuerza máxima que puede ejercer el músculo, la velocidad de contracción se hace cero y no se produce ninguna contracción, a pesar de la activación de la fibra muscular.
Un músculo esquelético se contrae muy rápido cuando no tiene ninguna carga, y va disminuyendo su velocidad conforme la carga va a aumentando. cuando la carga ha aumentado hasta la fuerza máxima que puede ejercer el músculo, la velocidad de contracción se hace cero y no se produce ninguna contracción, a pesar de la activación de la fibra muscular.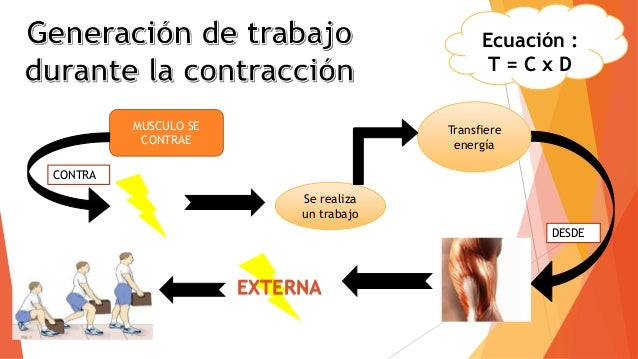 Cuando un músculo se contrae contra una carga realiza un trabajo. Esto significa que se transfiere energía desde el músculo hasta la carga externa para levantar un objeto hasta una mayor altura o para superar la resistencia al movimiento. El trabajo se define mediante la siguiente ecuación:
Cuando un músculo se contrae contra una carga realiza un trabajo. Esto significa que se transfiere energía desde el músculo hasta la carga externa para levantar un objeto hasta una mayor altura o para superar la resistencia al movimiento. El trabajo se define mediante la siguiente ecuación: La mioglobina se combina con el oxígeno y lo almacena hasta que sea necesario; esto también acelera mucho el transporte de oxígeno hacia las mitocondrias. La mioglobina da al músculo lento un aspecto rojizo y el nombre de músculo rojo.
La mioglobina se combina con el oxígeno y lo almacena hasta que sea necesario; esto también acelera mucho el transporte de oxígeno hacia las mitocondrias. La mioglobina da al músculo lento un aspecto rojizo y el nombre de músculo rojo.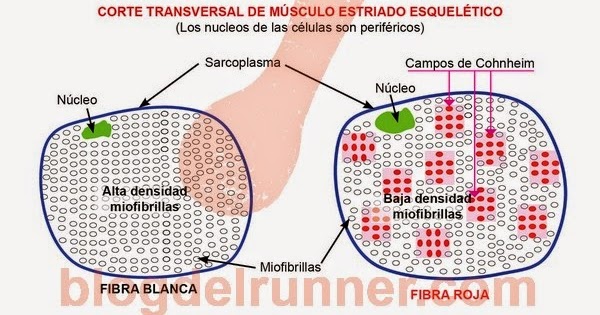
 Todas las fibras musculares que son inervadas por una única fibra nerviosa se denominan unidad motora. Los músculos pequeños que reaccionan rápidamente y cuyo control debe ser exacto tienen más fibras nerviosas para menos fibras musculares; en cambio, los músculos grandes que no precisan un control fino, como el músculo sóleo, pueden tener varios centenares de fibras musculares en una unidad motora.
Todas las fibras musculares que son inervadas por una única fibra nerviosa se denominan unidad motora. Los músculos pequeños que reaccionan rápidamente y cuyo control debe ser exacto tienen más fibras nerviosas para menos fibras musculares; en cambio, los músculos grandes que no precisan un control fino, como el músculo sóleo, pueden tener varios centenares de fibras musculares en una unidad motora.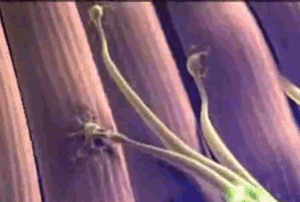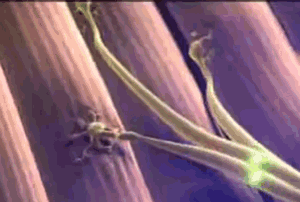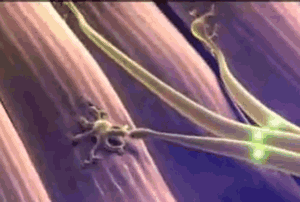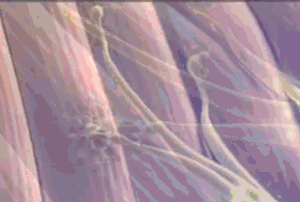 Las fibras musculares de todas las unidades motoras no están agrupadas entre sí en el músculo, sino que se juntan en otras formando microfascículos de 3 a 15 fibras. Esta interdigitación permite que las unidades motoras separadas se contraigan cooperando entre sí y no como segmentos totalmente individuales.
Las fibras musculares de todas las unidades motoras no están agrupadas entre sí en el músculo, sino que se juntan en otras formando microfascículos de 3 a 15 fibras. Esta interdigitación permite que las unidades motoras separadas se contraigan cooperando entre sí y no como segmentos totalmente individuales. El efecto de la escalera (Treppe): Cuando un músculo comienza a contraerse después de un período de reposo prolongado, su fuerza de contracción inicial puede ser tan pequeña, es decir, la fuerza de la contracción aumenta hasta una meseta, un fenómeno que se denomina efecto de la escalera o Treppe.
El efecto de la escalera (Treppe): Cuando un músculo comienza a contraerse después de un período de reposo prolongado, su fuerza de contracción inicial puede ser tan pequeña, es decir, la fuerza de la contracción aumenta hasta una meseta, un fenómeno que se denomina efecto de la escalera o Treppe.
 Tono muscular: son los músculos que se encuentran «relajados», pero tiene una ligera contracción.
Tono muscular: son los músculos que se encuentran «relajados», pero tiene una ligera contracción.



 La membrana celular está formada en su mayoría por una bicapa lipídica, aunque también contiene varias moléculas proteicas insertadas en los lípidos, muchas de ellas penetran en todo el grosor de la membrana. La bicapa lipídica no es miscible con el líquido extracelular ni con el líquido intracelular. Por tanto, constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular.
La membrana celular está formada en su mayoría por una bicapa lipídica, aunque también contiene varias moléculas proteicas insertadas en los lípidos, muchas de ellas penetran en todo el grosor de la membrana. La bicapa lipídica no es miscible con el líquido extracelular ni con el líquido intracelular. Por tanto, constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular. Otras, denominadas proteínas transportadoras, se unen a las moléculas o iones que se van a transportar produciendose cambios conformacionales de las moléculas de la proteína desplazan después las sustancias a través de los intersticios de la proteína hasta el otro lado de la membrana. Tanto las proteínas de los canales como las proteínas transportadoras habitualmente son muy selectivas para los tipos de moléculas o de iones que pueden atravesar la membrana.
Otras, denominadas proteínas transportadoras, se unen a las moléculas o iones que se van a transportar produciendose cambios conformacionales de las moléculas de la proteína desplazan después las sustancias a través de los intersticios de la proteína hasta el otro lado de la membrana. Tanto las proteínas de los canales como las proteínas transportadoras habitualmente son muy selectivas para los tipos de moléculas o de iones que pueden atravesar la membrana.
 Entre las sustancias más importantes que atraviesan las membranas celulares mediante difusión facilitada están la glucosay la mayor parte de los aminoácidos. Este también es el principal mecanismo mediante el cual la insulina controla la utilización de glucosa por el cuerpo.
Entre las sustancias más importantes que atraviesan las membranas celulares mediante difusión facilitada están la glucosay la mayor parte de los aminoácidos. Este también es el principal mecanismo mediante el cual la insulina controla la utilización de glucosa por el cuerpo. Por ejemplo, los poros proteicos denominados acuaporinas o canales de agua permiten el rápido paso de agua a través de las membranas celu-lares pero impiden el de otras moléculas.
Por ejemplo, los poros proteicos denominados acuaporinas o canales de agua permiten el rápido paso de agua a través de las membranas celu-lares pero impiden el de otras moléculas. Muchos de los canales proteicos son muy selectivos para el transporte de uno o más iones o moléculas específicos. Esto se debe a las características del propio canal, como su diámetro, su forma y la naturaleza de las cargas eléctricas y enlaces químicos que están situados a lo largo de sus superficies internas.
Muchos de los canales proteicos son muy selectivos para el transporte de uno o más iones o moléculas específicos. Esto se debe a las características del propio canal, como su diámetro, su forma y la naturaleza de las cargas eléctricas y enlaces químicos que están situados a lo largo de sus superficies internas.



 Transporte activo secundario: la energía procede secundariamente de la energía que se ha almacenado en forma de diferencias de concentración iónica de sustancias moleculares o iónicas secundarias entre los dos lados de una membrana celular, que se generó originalmente mediante transporte activo primario.
Transporte activo secundario: la energía procede secundariamente de la energía que se ha almacenado en forma de diferencias de concentración iónica de sustancias moleculares o iónicas secundarias entre los dos lados de una membrana celular, que se generó originalmente mediante transporte activo primario.

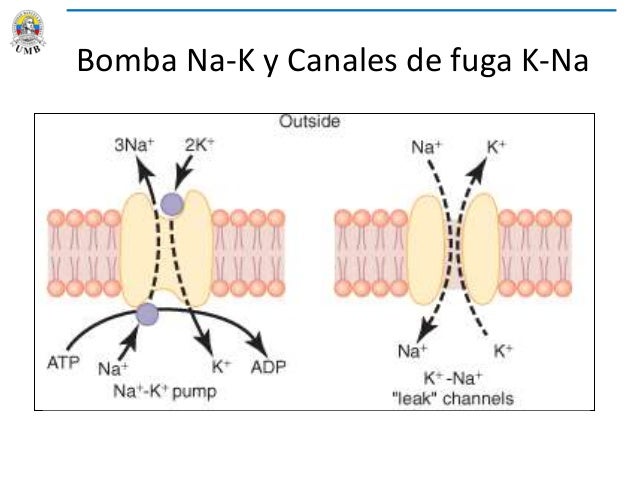 Una proteína del canal, a veces denominada dominio de poros en tándem, canal de potasio o canal de fuga de potasio también puede dejar que se pierdan algunos iones de sodio, sin embargo, siempre van a ser más permeables al K; ésta diferencia de permeabilidad es un factor importante para determinar el nivel del potencial de membrana en reposo normal.
Una proteína del canal, a veces denominada dominio de poros en tándem, canal de potasio o canal de fuga de potasio también puede dejar que se pierdan algunos iones de sodio, sin embargo, siempre van a ser más permeables al K; ésta diferencia de permeabilidad es un factor importante para determinar el nivel del potencial de membrana en reposo normal.
 Fase de reposo: Es el potencial de membrana en reposo antes del comienzo del potencial de acción, la membrana está polarizada durante esta fase debido al potencial de membrana negativo de –90 mV que posee.
Fase de reposo: Es el potencial de membrana en reposo antes del comienzo del potencial de acción, la membrana está polarizada durante esta fase debido al potencial de membrana negativo de –90 mV que posee.
 Activación: Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje ( entre –70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta pudiendo así pasar los iones sodio.
Activación: Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje ( entre –70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta pudiendo así pasar los iones sodio. Durante el estado de reposo la compuerta del canal de potasio está cerrada, lo que impide que los iones potasio pasen a través de este canal hacia el exterior. Cuando el potencial de membrana aumenta desde –90 mV hacia cero, este voltaje produce que la compuerta se abra y permite el aumento de la difusión de potasio hacia fuera a través del canal, pero debido a que dilata un poco la apertura de los canales de potasio, en su mayor parte, se abren al mismo tiempo que se están cerrando los canales de sodio debido a su inactivación.
Durante el estado de reposo la compuerta del canal de potasio está cerrada, lo que impide que los iones potasio pasen a través de este canal hacia el exterior. Cuando el potencial de membrana aumenta desde –90 mV hacia cero, este voltaje produce que la compuerta se abra y permite el aumento de la difusión de potasio hacia fuera a través del canal, pero debido a que dilata un poco la apertura de los canales de potasio, en su mayor parte, se abren al mismo tiempo que se están cerrando los canales de sodio debido a su inactivación.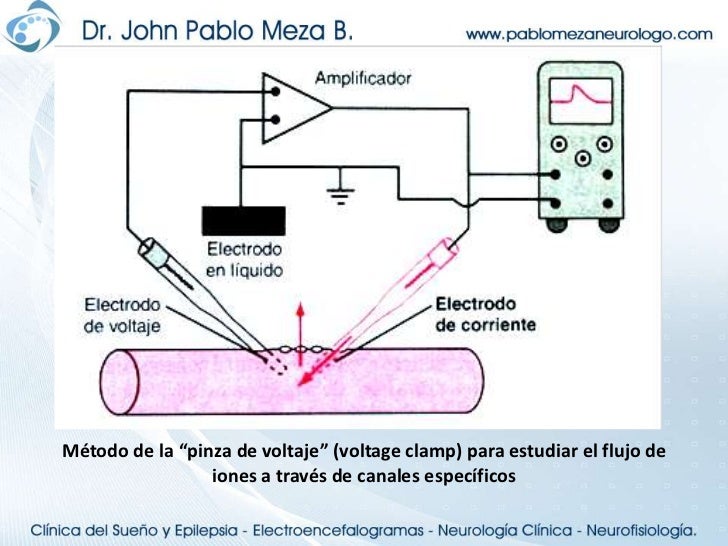 Es un método experimental que se utiliza para medir el flujo de iones a través de los diferentes canales. Cuando se utiliza este aparato se insertan dos electrodos en la fibra nerviosa. Uno de ellos sirve para medir el voltaje del potencial de membrana y el otro para conducir corriente eléctrica hacia el interior o el exterior de la fibra nerviosa.
Es un método experimental que se utiliza para medir el flujo de iones a través de los diferentes canales. Cuando se utiliza este aparato se insertan dos electrodos en la fibra nerviosa. Uno de ellos sirve para medir el voltaje del potencial de membrana y el otro para conducir corriente eléctrica hacia el interior o el exterior de la fibra nerviosa. Una función importante de los canales de iones calcio activados por voltaje consiste en su contribución a la fase de despolarización en el potencial de acción en algunas células. Se denominan canales lentos, en contraposición a los canales de sodio, que se denominan canales rápidos. Por tanto, la apertura de los canales de calcio proporciona una despolarización más sostenida, mientras que los canales de sodio desempeñan un papel clave en la iniciación de los potenciales de acción. Hay abundantes canales de calcio tanto en el músculo cardíaco como el músculo liso.
Una función importante de los canales de iones calcio activados por voltaje consiste en su contribución a la fase de despolarización en el potencial de acción en algunas células. Se denominan canales lentos, en contraposición a los canales de sodio, que se denominan canales rápidos. Por tanto, la apertura de los canales de calcio proporciona una despolarización más sostenida, mientras que los canales de sodio desempeñan un papel clave en la iniciación de los potenciales de acción. Hay abundantes canales de calcio tanto en el músculo cardíaco como el músculo liso.
 No se puede producir un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen. Esto se produce cuando el número de iones Na+ que entran en la fibra supera al número de iones K+ que salen de la misma. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Se dice que el nivel de –65 mV es el umbral para la estimulación.
No se puede producir un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen. Esto se produce cuando el número de iones Na+ que entran en la fibra supera al número de iones K+ que salen de la misma. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Se dice que el nivel de –65 mV es el umbral para la estimulación.

 En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y sólo después comienza la repolarización. Este tipo de potencial de acción se produce en las fibras musculares cardíacas.
En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y sólo después comienza la repolarización. Este tipo de potencial de acción se produce en las fibras musculares cardíacas.



 El núcleo central de la fibra es el axón, y la membrana del axón es la membrana que realmente conduce el potencial de acción. El axón contiene en su centro el axoplasma, que es un líquido intracelular viscoso. Alrededor del axón hay una vaina de mielina, que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier.
El núcleo central de la fibra es el axón, y la membrana del axón es la membrana que realmente conduce el potencial de acción. El axón contiene en su centro el axoplasma, que es un líquido intracelular viscoso. Alrededor del axón hay una vaina de mielina, que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier. Los iones no pueden fluir a través de las gruesas vainas de mielina de los nervios mielinizados, pueden fluir fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen sólo en los nódulos. Esto se denomina conducción saltatoria. Es decir, la corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro.
Los iones no pueden fluir a través de las gruesas vainas de mielina de los nervios mielinizados, pueden fluir fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen sólo en los nódulos. Esto se denomina conducción saltatoria. Es decir, la corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro. La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan sólo 0,25 m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan sólo 0,25 m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
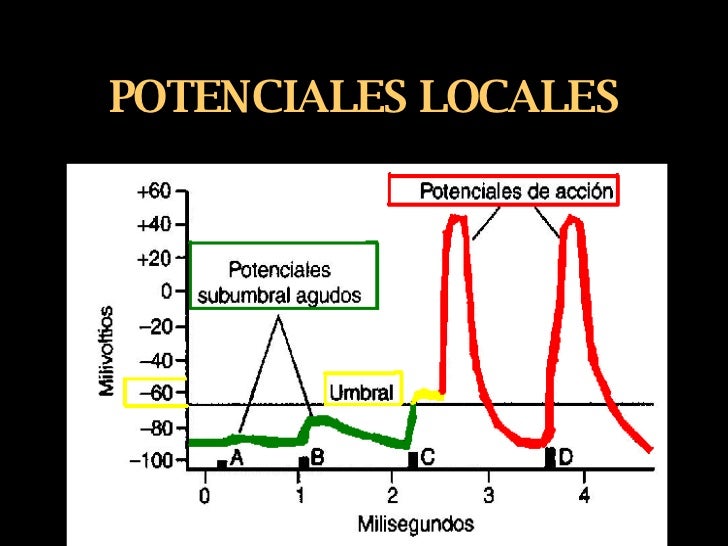 Los cambios locales de potencial se denominan potenciales locales agudos y, cuando no pueden generarun potencial de acción, se denominan potenciales subliminales agudos. Cuando el potencial local apenas alcanza el nivel necesario para generar un potencial de acción, se denomina nivel liminar (umbral).
Los cambios locales de potencial se denominan potenciales locales agudos y, cuando no pueden generarun potencial de acción, se denominan potenciales subliminales agudos. Cuando el potencial local apenas alcanza el nivel necesario para generar un potencial de acción, se denomina nivel liminar (umbral). Los factores estabilizadores de la membrana son aquellos que pueden reducir la excitabilidad, por ejemplo, el ion calcio es un estabilizador.
Los factores estabilizadores de la membrana son aquellos que pueden reducir la excitabilidad, por ejemplo, el ion calcio es un estabilizador.

