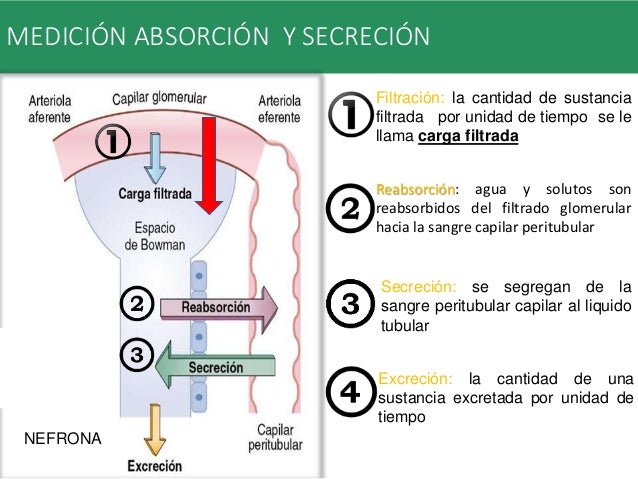
Reabsorción y secreción tubular renal
 A medida que el filtrado glomerular pasa por los túbulos renales, fluye de forma secuencial a través de sus diferentes partes (el túbulo proximal, el asa de Henle, el túbulo distal, el túbulo colector y, finalmente, el conducto colector) antes de eliminarse por la orina.
A medida que el filtrado glomerular pasa por los túbulos renales, fluye de forma secuencial a través de sus diferentes partes (el túbulo proximal, el asa de Henle, el túbulo distal, el túbulo colector y, finalmente, el conducto colector) antes de eliminarse por la orina.
La reabsorción tubular es cuantitativamente importante y altamente selectiva
Filtración = Filtrado glomerular
× Concentración plasmática
 Los procesos de la filtración glomerular y de la reabsorción tubular son cuantitativamente muy intensos en comparación con la excreción urinaria de muchas sustancias. Esto significa que en un pequeño cambio en la filtración glomerular o en la reabsorción tubular podría causar un
Los procesos de la filtración glomerular y de la reabsorción tubular son cuantitativamente muy intensos en comparación con la excreción urinaria de muchas sustancias. Esto significa que en un pequeño cambio en la filtración glomerular o en la reabsorción tubular podría causar un
cambio relativamente importante en la excreción urinaria.
 A diferencia de la filtración glomerular, que carece relativamente de selectividad (prácticamente todos los solutos del plasma se filtran salvo las proteínas del plasma o las sustancias unidas a ellas), la reabsorción tubular es muy selectiva. Algunas sustancias, como la glucosa y los aminoácidos, se reabsorben del todo en los túbulos, por lo que su excreción urinaria es prácticamente nula.
A diferencia de la filtración glomerular, que carece relativamente de selectividad (prácticamente todos los solutos del plasma se filtran salvo las proteínas del plasma o las sustancias unidas a ellas), la reabsorción tubular es muy selectiva. Algunas sustancias, como la glucosa y los aminoácidos, se reabsorben del todo en los túbulos, por lo que su excreción urinaria es prácticamente nula.
La reabsorción tubular comprende mecanismos pasivos y activos
Para que una sustancia se reabsorba, primero debe ser transportada: 1) a través de las membranas del epitelio tubular hasta el líquido intersticial renal. 2) a través de la membrana capilar peritubular hasta la sangre.
Transporte activo
El transporte activo puede mover un soluto en contra de un gradiente electroquímico y para ello precisa energía del metabolismo.


La reabsorción de glucosa por el túbulo renal es un ejemplo de transporte activo secundario.
Los solutos pueden transportarse a través de las células epiteliales o entre las células.
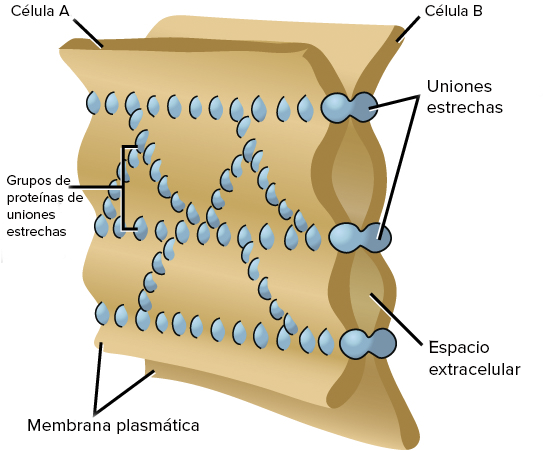 Las células tubulares renales, al igual que otras células epiteliales, se mantienen juntas por medio de uniones estrechas. Los espacios intercelulares laterales están situados por detrás de estas uniones estrechas y separan las células epiteliales del túbulo.
Las células tubulares renales, al igual que otras células epiteliales, se mantienen juntas por medio de uniones estrechas. Los espacios intercelulares laterales están situados por detrás de estas uniones estrechas y separan las células epiteliales del túbulo.
Los solutos pueden reabsorberse o secretarse a través de las células por vía transcelular o entre las células moviéndose a través de las uniones estrechas y los espacios intercelulares siguiendo la vía paracelular.

El transporte activo primario a través de la membrana tubular está acoplado a la hidrólisis del ATP. La importancia especial del trasporte activo primario es que puede mover los solutos en contra de un gradiente electroquímico. La energía necesaria para este transporte activo procede de la hidrólisis del ATP que realiza la ATPasa unida a la membrana; la ATPasa es también un componente del mecanismo de transporte que liga y mueve solutos a través de las membranas celulares. Los transportadores activos primarios en los riñones que conocemos son la ATPasa sodiopotasio, la ATPasa hidrógeno, la ATPasa hidrógenopotasio y la ATPasa calcio.
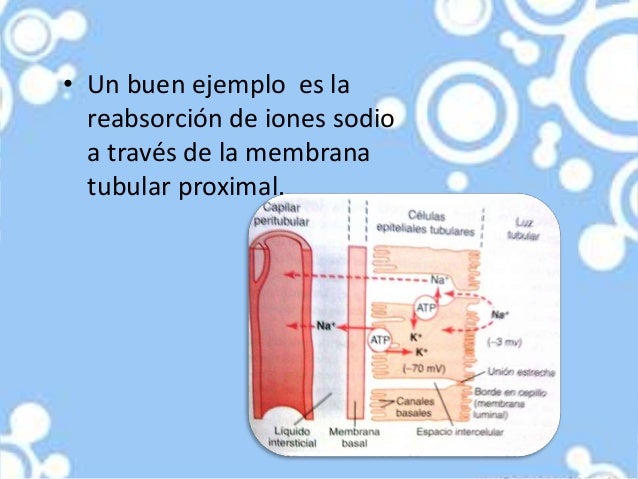
El funcionamiento de esta bomba de iones mantiene una concentración intracelular de sodio baja y una concentración intracelular de potasio alta y genera una carga negativa neta de unos –70 mV dentro
de la célula.
La reabsorción activa del sodio mediante la ATPasa sodio-potasio tiene lugar en la mayor parte del túbulo.
Así pues, la reabsorción neta de los iones sodio desde la luz tubular hacia la sangre supone al menos tres pasos:
1. El sodio se difunde a través de la membrana luminal (también llamada membrana apical) al interior de la célula siguiendo un gradiente electroquímico creado por la bomba ATPasa sodiopotasio.
2. El sodio es transportado a través de la membrana basolateral contra un gradiente electroquímico por la acción de la bomba ATPasa sodiopotasio.
3. El sodio, el agua y otras sustancias se reabsorben del líquido intersticial hacia los capilares peritubulares por ultrafiltración, un proceso pasivo gobernado por gradientes de presión hidrostática y coloidosmótica.
Reabsorción activa secundaria a través de la membrana tubular.
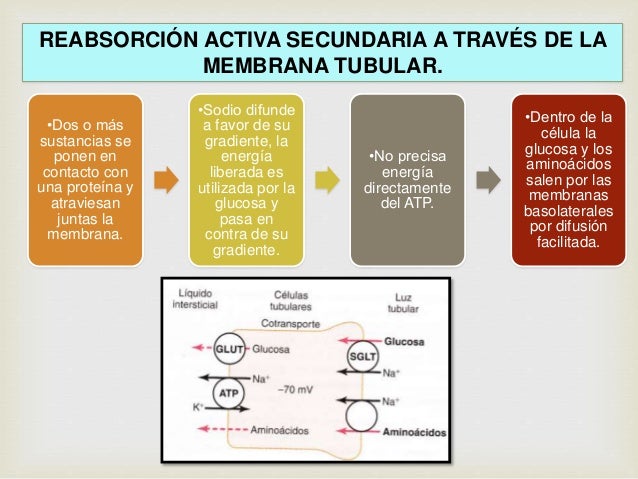
Los cotransportadores de glucosa y sodio (SGLT2 y SGLT1) están situados en el borde en cepillo de las células tubulares proximales y llevan glucosa al citoplasma celular en contra de un gradiente de concentración, como se ha descrito antes. Aproximadamente el 90% de la glucosa filtrada es reabsorbido por SGLT2 en la primera parte del túbulo proximal y el 10% residual es transportado por SGLT1 en los segmentos posteriores del túbulo proximal. En el lado basolateral de la membrana, la glucosa se difunde fuera de la célula a los espacios intersticiales con la ayuda de transportadores de glucosa: GLUT2, en el segmento S1 y GLUT1 en la última parte (segmento S3) del túbulo proximal.
Secreción activa secundaria hacia los túbulos. Algunas sustancias se secretan en los túbulos mediante un transporte activo secundario. Esto supone a menudo un contratransporte de la sustancia junto a iones sodio.
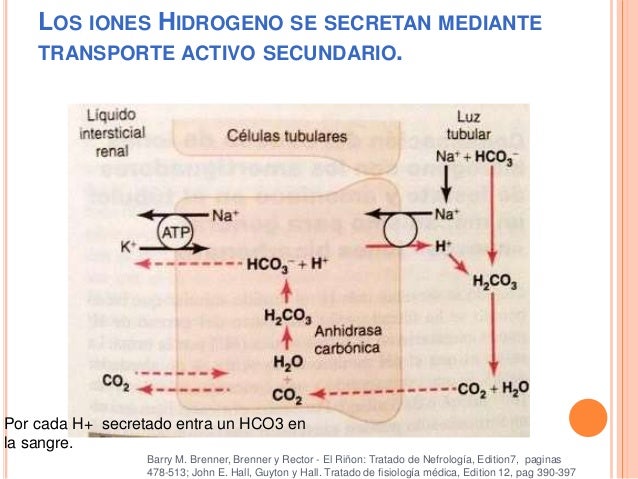
Un ejemplo de contratransporte, es la secreción activa de iones hidrógeno acoplada a la reabsorción de sodio en la membrana luminal del túbulo proximal.
Transporte máximo de sustancias que se reabsorben de forma activa.
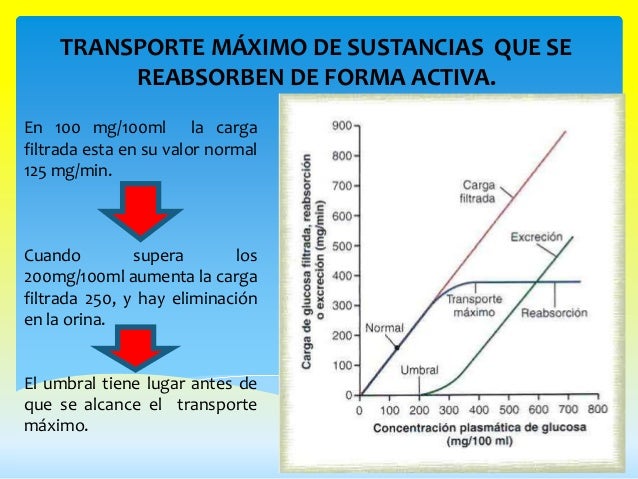
Para la mayoría de las sustancias que se reabsorben o excretan activamente hay un límite en la intensidad con la que pueden transportarse, denominado a menudo transporte máximo.

Sustancias que se transportan de forma activa pero no exhiben transporte máximo.
La razón de que solutos con trasporte activo muestren a menudo un transporte máximo es que el sistema transportador se satura a medida que la carga tubular aumenta. Algunas sustancias que se reabsorben de forma pasiva no muestran un transporte máximo porque la intensidad de su transporte está determinada por otros factores.

La reabsorción pasiva del agua mediante ósmosis está acoplada sobre todo a la reabsorción de sodio
Cuando los solutos se transportan fuera del túbulo mediante un transporte activo primario o secundario, sus concentraciones tienden a reducirse dentro del túbulo y a aumentaren el intersticio renal. Esto crea una diferencia de concentración que produce la ósmosis del agua en la misma dirección que la de los solutos que se transportan, desde la luz tubular hacia el intersticio renal.
Una gran parte del flujo osmótico de agua en los túbulos proximales se produce a través de las también conocidas como uniones estrechas que hay entre las células epiteliales y
a través de las propias células.
 La reabsorción activa de sodio está muy bien acoplada a la reabsorción pasiva de cloro a través de un potencial eléctrico y un gradiente de concentración de cloro.
La reabsorción activa de sodio está muy bien acoplada a la reabsorción pasiva de cloro a través de un potencial eléctrico y un gradiente de concentración de cloro.
Los iones cloro pueden reabsorberse también mediante un transporte activo secundario. El más importante de los procesos activos secundarios para la reabsorción del cloro consiste en el cotransporte del cloro con el sodio a través de la membrana luminal.
La urea también se reabsorbe de forma pasiva del túbulo, pero en un grado mucho menor que los iones cloro. A medida que el agua se reabsorbe de los túbulos (por ósmosis acoplada a la reabsorción de sodio), la concentración de urea en la luz tubular aumenta.
Reabsorción y secreción a lo largo de diferentes partes de la nefrona
Reabsorción en el túbulo proximal
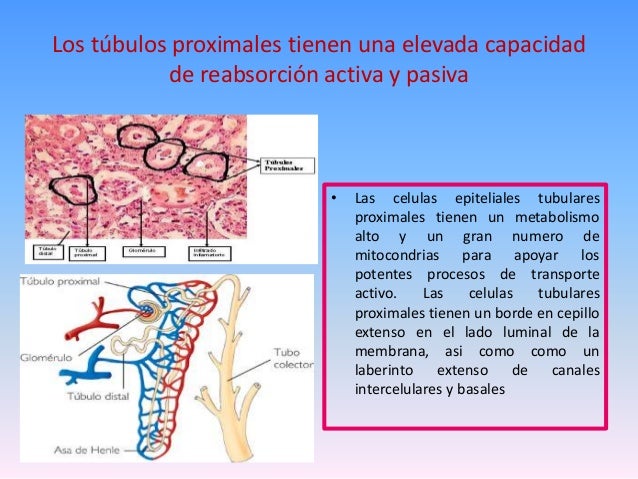
Alrededor del 65% de la carga filtrada de sodio y agua y algo menos del cloro filtrado se reabsorbe normalmente en el túbulo proximal antes de que el filtrado alcance el asa de Henle.
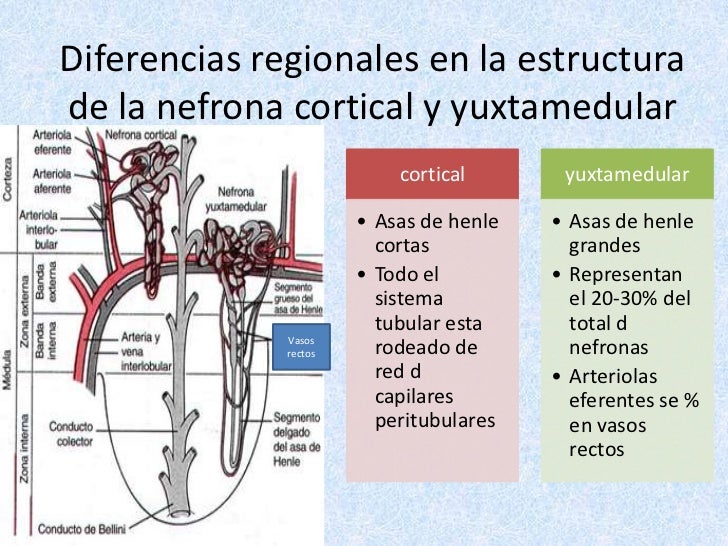
Transporte de solutos y agua en el asa de Henle
El asa de Henle consta de tres segmentos con funciones diferentes: el segmento descendente fino, el segmento ascendente fino y el segmento ascendente grueso.
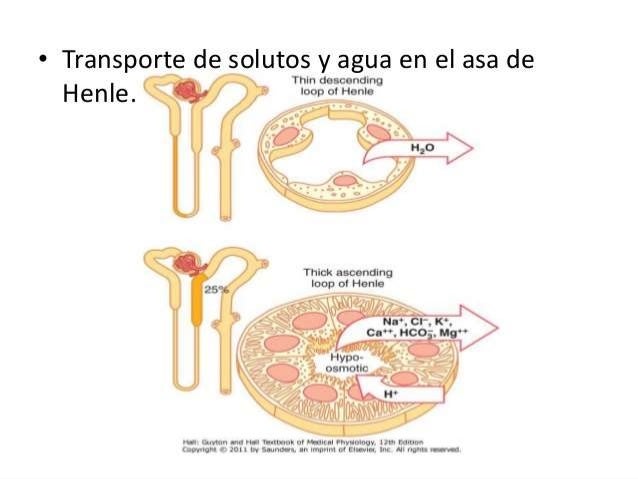 La parte descendente del segmento fino es muy permeable al agua y moderadamente a la mayoría de los solutos, incluidos la urea y el sodio. La función de este segmento de la nefrona es sobre todo permitir la difusión simple de las sustancias a través de sus paredes. La rama ascendente, incluidas las porciones fina y gruesa, es casi impermeable al agua, una característica que es importante para concentrar la orina.
La parte descendente del segmento fino es muy permeable al agua y moderadamente a la mayoría de los solutos, incluidos la urea y el sodio. La función de este segmento de la nefrona es sobre todo permitir la difusión simple de las sustancias a través de sus paredes. La rama ascendente, incluidas las porciones fina y gruesa, es casi impermeable al agua, una característica que es importante para concentrar la orina.
El segmento grueso del asa de Henle, que comienza en la mitad de la rama ascendente, tiene células epiteliales gruesas que tienen una elevada actividad metabólica y son capaces de una reabsorción activa del sodio, el cloro y el potasio. También se reabsorben cantidades considerables de otros iones, como calcio, bicarbonato y magnesio, en la rama ascendente gruesa del asa de Henle. El segmento fino de la rama ascendente tiene un capacidad de reabsorción mucho menor que el segmento grueso y la rama descendente fina no reabsorbe cantidades significativas de ninguno de estos solutos.
Túbulo distal
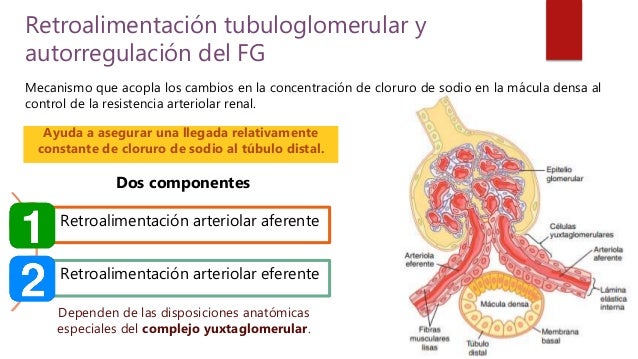
 El segmento grueso de la rama ascendente del asa de Henle se vacía en el túbulo distal. La porción inicial del túbulo distal conforma la mácula densa, un grupo de células epiteliales densamente empaquetadas que es parte del complejo yuxtaglomerular que proporciona un control de retroalimentación del FG y del flujo sanguíneo en esta misma nefrona.
El segmento grueso de la rama ascendente del asa de Henle se vacía en el túbulo distal. La porción inicial del túbulo distal conforma la mácula densa, un grupo de células epiteliales densamente empaquetadas que es parte del complejo yuxtaglomerular que proporciona un control de retroalimentación del FG y del flujo sanguíneo en esta misma nefrona.
El segmento diluyente reabsorbe con avidez la mayoría de los iones, incluidos el sodio, el potasio y el cloro, pero es casi totalmente impermeable al agua y a la urea.
Porción final del túbulo distal y túbulo colector cortical
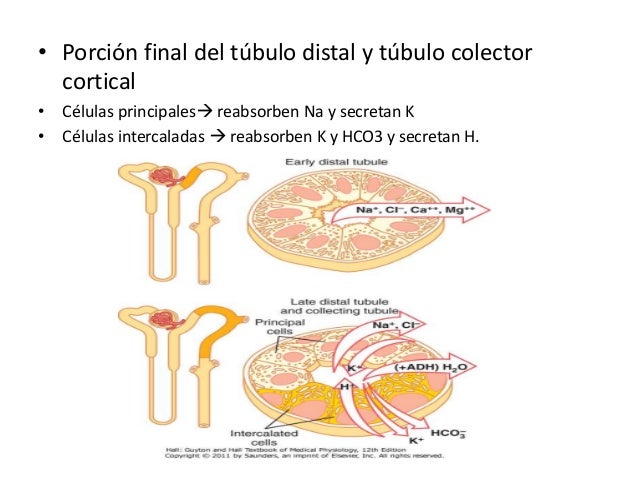
Las células principales reabsorben sodio y secretan potasio.
La reabsorción de sodio y la secreción de potasio por las células principales depende de la actividad de la bomba ATPasa sodiopotasio presente en la membrana basolateral de cada célula.
Las células principales son los primeros lugares de acción de los diuréticos ahorradores de potasio, como espironolactona, eplerenona, amilorida y triamtereno. Los antagonistas de los receptores de espironolactona y eplerenona compiten con la aldosterona por sus receptores en las células principales y por tanto inhiben los efectos estimuladores de esta hormona sobre la reabsorción de sodio y la secreción de potasio. La amilorida y el triamtereno son bloqueantes de los canales del sodio que inhiben directamente la entrada del sodio en los canales del sodio de las membranas luminales y así reducen la cantidad de sodio que puede transportarse a través de las membranas basolaterales por medio de la bomba ATPasa sodiopotasio.

Conducto colector medular
Aunque los conductos colectores medulares reabsorben menos del 10% del agua y del sodio filtrados, son el lugar final de procesamiento de la orina.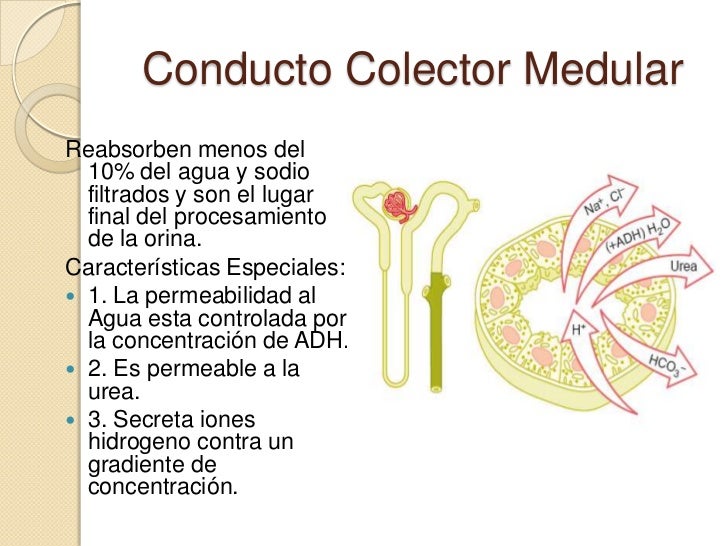
Regulación de la reabsorción tubular
Una característica importante de la reabsorción tubular es que la reabsorción de algunos solutos puede regularse independientemente de la de otros, en especial mediante mecanismos de control hormonal.
Equilibrio glomerulotubular: la capacidad de los túbulos de aumentar la reabsorción en respuesta a un incremento de la carga tubular
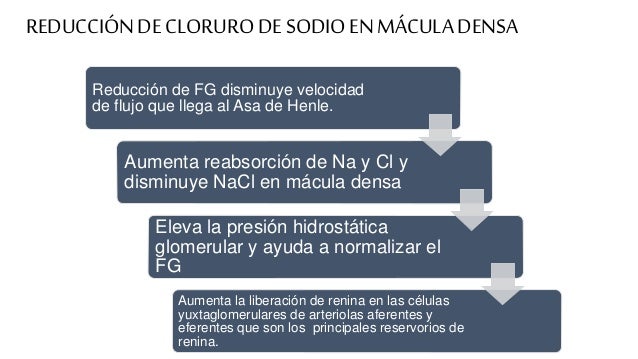
Uno de los mecanismos más básicos de control de la reabsorción tubular es la capacidad intrínseca de los túbulos de aumentar su reabsorción en respuesta a una mayor carga tubular (un aumento del flujo tubular). Este fenómeno se denomina equilibrio glomerulotubular.
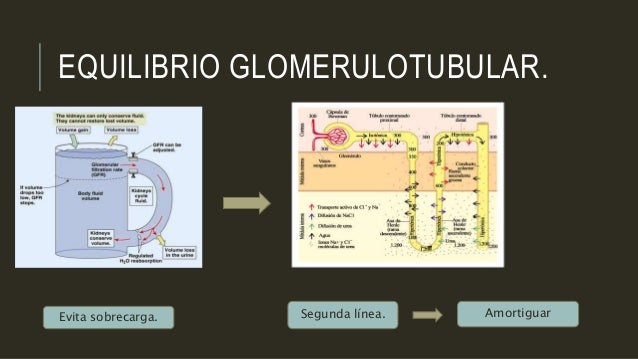
El equilibrio glomerulotubular ayuda a evitar sobrecargas en segmentos del túbulo distal cuando el FG aumenta.
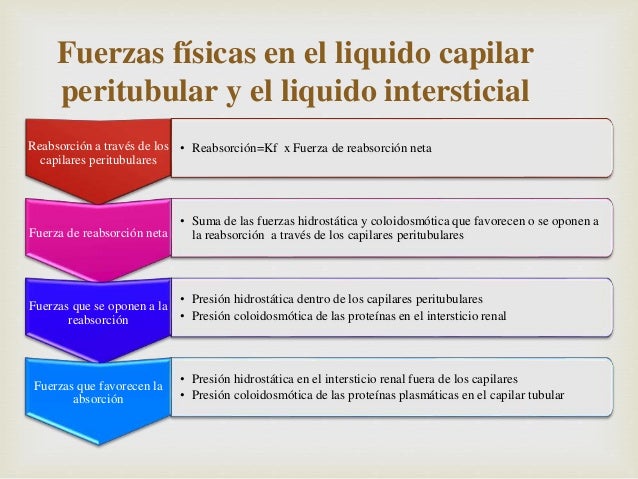
Fuerzas físicas en el líquido capilar peritubular y el líquido intersticial
Las fuerzas hidrostática y coloidosmótica gobiernan el grado de reabsorción a través de los capilares peritubulares, a la vez que controlan la filtración en los capilares glomerulares.
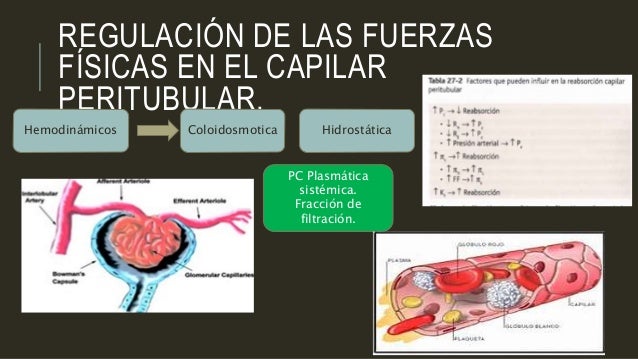
Regulación de las fuerzas físicas en el capilar peritubular.

Presiones hidrostática y coloidosmótica en el intersticio renal.

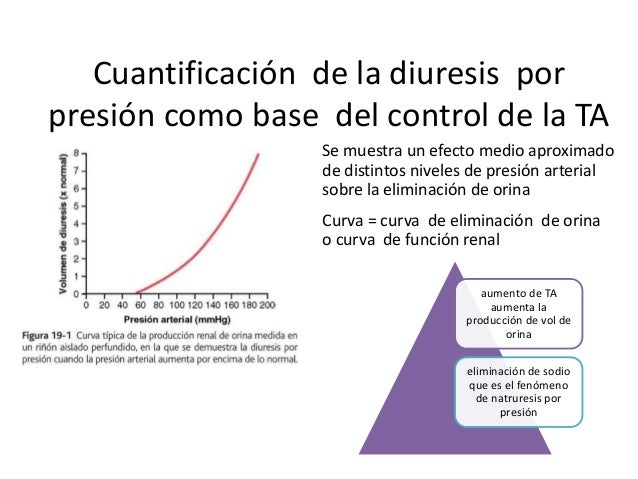
Efecto de la presión arterial sobre la diuresis: presión-natriuresis y presión-diuresis
Incluso pequeños incrementos en la presión arterial pueden provocar aumentos en la excreción urinaria de sodio y agua, fenómenos que se conocen como natriuresis por presión y diuresis por presión.

Control hormonal de la reabsorción tubular
La aldosterona aumenta la reabsorción de sodio y estimula la secreción de potasio.
 La aldosterona, que secretan las células de la glomerulosa de la corteza suprarrenal, es un regulador importante de la reabsorción de sodio y la secreción de potasio en los túbulos renales. Un lugar de acción tubular renal importante de la aldosterona son las células principales del túbulo colector cortical. El mecanismo por el cual la aldosterona aumenta la reabsorción de sodio mientras incrementa a la vez la secreción de potasio es estimulando la bomba ATPasa sodio-potasio en el lado basolateral de la membrana del túbulo colector cortical. La aldosterona también aumenta la permeabilidad al sodio del lado luminal de la membrana.
La aldosterona, que secretan las células de la glomerulosa de la corteza suprarrenal, es un regulador importante de la reabsorción de sodio y la secreción de potasio en los túbulos renales. Un lugar de acción tubular renal importante de la aldosterona son las células principales del túbulo colector cortical. El mecanismo por el cual la aldosterona aumenta la reabsorción de sodio mientras incrementa a la vez la secreción de potasio es estimulando la bomba ATPasa sodio-potasio en el lado basolateral de la membrana del túbulo colector cortical. La aldosterona también aumenta la permeabilidad al sodio del lado luminal de la membrana.
Los estímulos más importantes para la aldosterona son:
1) aumento de la concentración extracelular de potasio.
2) aumento de los niveles de angiotensina II.
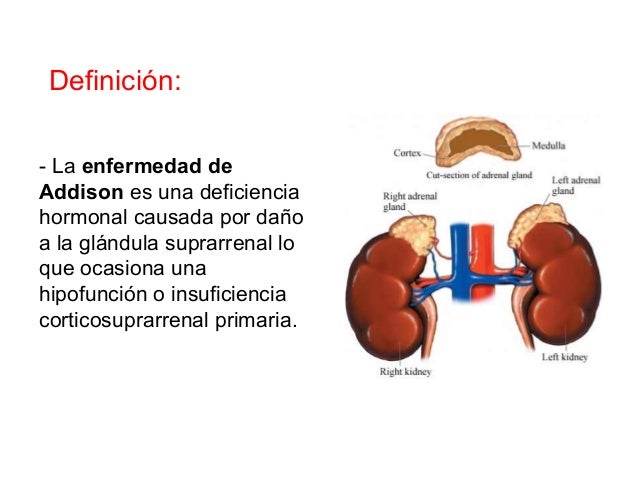 Sin aldosterona, como ocurre en la destrucción o mala función de la glándula supra-rrenal (enfermedad de Addison), hay una pérdida acentuada de sodio y una acumulación de potasio en el organismo.
Sin aldosterona, como ocurre en la destrucción o mala función de la glándula supra-rrenal (enfermedad de Addison), hay una pérdida acentuada de sodio y una acumulación de potasio en el organismo.


El exceso de secreción de aldosterona, como ocurre en los pacientes con tumores suprarrenales (síndrome de Conn) se acompaña de una retención de sodio y una disminución de potasio en plasma debida, en parte, a una excesiva secreción de potasio por los riñones.
La angiotensina II aumenta la reabsorción de sodio y de agua.
La angiotensina II es quizás la hormona ahorradora de sodio más potente del organismo. La mayor formación de angiotensina II ayuda a normalizar la presión arterial y el volumen extracelular al aumentar la reabsorción de sodio y agua en los túbulos renales a través de tres efectos principales:
1. La angiotensina II estimula la secreción de aldosterona, lo que a su vez aumenta la reabsorción de sodio.
2. La angiotensina II contrae las arteriolas eferentes, lo que tiene dos efectos sobre la dinámica capilar peritubular que aumentan el sodio y el agua.
- La constricción arteriolar reduce la presión hidrostática capilar peritubular, lo que aumenta la reabsorción tubular neta, en especial en los túbulos proximales.
- La constricción arteriolar eferente, al reducir el flujo sanguíneo, aumenta la fracción de filtración en el glomérulo y también la concentración de proteínas y la presión coloidosmótica en los capilares peritubulares; esto incrementa la fuerza de reabsorción en los capilares peritubulares y la reabsorción tubular de sodio y agua.
3. 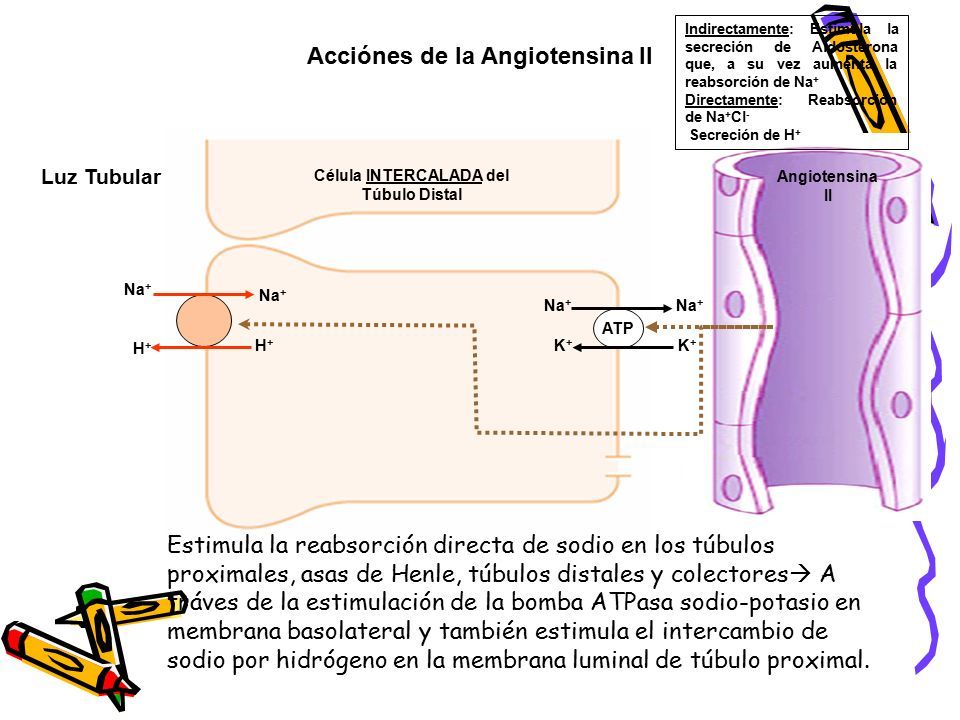 Uno de los efectos directos de la angiotensina II es estimular la bomba ATPasa sodiopotasio en la membrana basocelular de la célula epitelial tubular. Un segundo efecto es estimular el intercambio de sodio por hidrógeno en la membrana luminal, en especial en el túbulo proximal Un tercer efecto de la angiotensina II consiste en estimular el cotransporte de bicarbonatosodio en la membrana basolateral.
Uno de los efectos directos de la angiotensina II es estimular la bomba ATPasa sodiopotasio en la membrana basocelular de la célula epitelial tubular. Un segundo efecto es estimular el intercambio de sodio por hidrógeno en la membrana luminal, en especial en el túbulo proximal Un tercer efecto de la angiotensina II consiste en estimular el cotransporte de bicarbonatosodio en la membrana basolateral.
La ADH aumenta la reabsorción de agua. 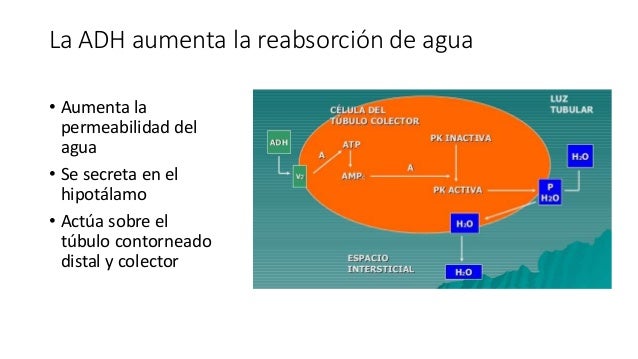
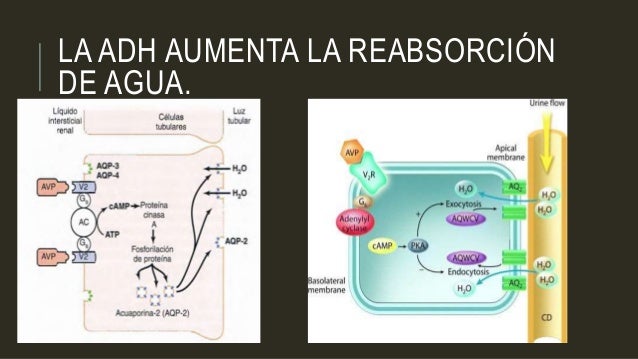 Este efecto ayuda al organismo a conservar el agua en circunstancias como la deshidratación. Sin ADH, la permeabilidad al agua de los túbulos distales y de los conductos colectores es baja, lo que hace que los riñones excreten grandes cantidades de orina diluida.
Este efecto ayuda al organismo a conservar el agua en circunstancias como la deshidratación. Sin ADH, la permeabilidad al agua de los túbulos distales y de los conductos colectores es baja, lo que hace que los riñones excreten grandes cantidades de orina diluida.
La hormona paratiroidea aumenta la reabsorción de calcio.
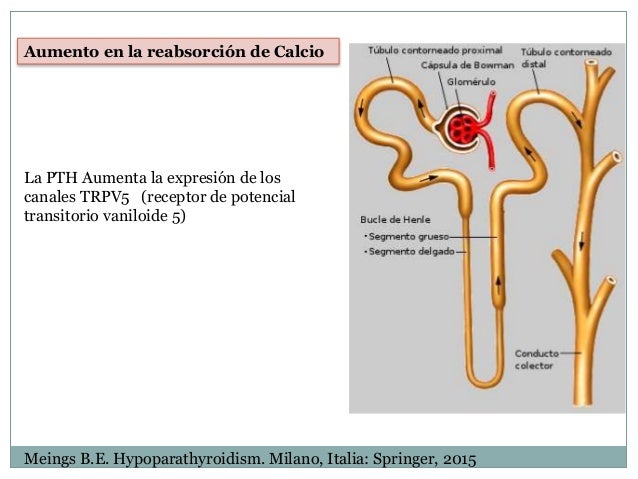 La hormona paratiroidea es una de las hormonas reguladoras del calcio más importantes del cuerpo. Su principal acción en los riñones es aumentar la reabsorción tubular de calcio, en especial en los túbulos distales y quizás también en las asas de Henle.
La hormona paratiroidea es una de las hormonas reguladoras del calcio más importantes del cuerpo. Su principal acción en los riñones es aumentar la reabsorción tubular de calcio, en especial en los túbulos distales y quizás también en las asas de Henle.
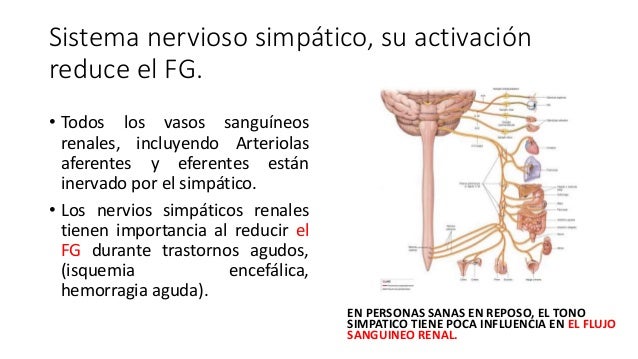
La activación del sistema nervioso simpático aumenta la reabsorción de sodio
La estimulación del sistema nervioso simpático aumenta la liberación de renina y la formación de angiotensina II, lo que contribuye al efecto global de aumento de la reabsorción tubular y reducción de la excreción renal de sodio.
Uso de los métodos de aclaramiento para cuantificar la función renal

BRENDA ALEJANDRA MARTÍNEZ FUENTES 1° B


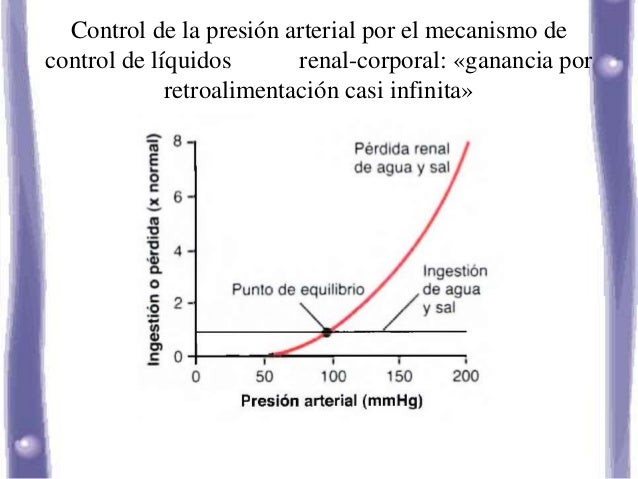
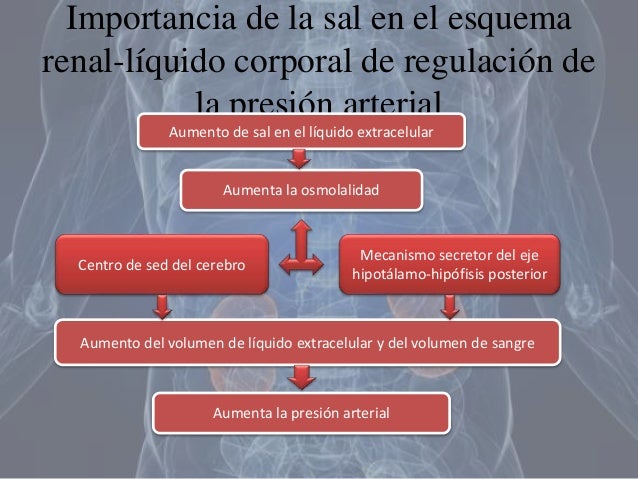
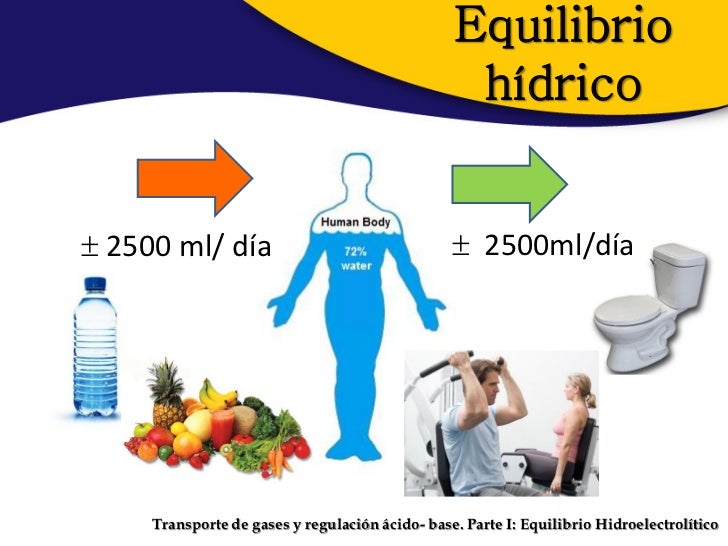 Para el mantenimiento de la homeostasis, la excreción de agua y electrólitos debe corresponderse de forma precisa con su ingreso. Si los ingresos superan a la excreción, la cantidad de esa sustancia en el cuerpo aumentará. Si la ingestión es menor que la excreción, la cantidad de esa sustancia en el cuerpo se reducirá.
Para el mantenimiento de la homeostasis, la excreción de agua y electrólitos debe corresponderse de forma precisa con su ingreso. Si los ingresos superan a la excreción, la cantidad de esa sustancia en el cuerpo aumentará. Si la ingestión es menor que la excreción, la cantidad de esa sustancia en el cuerpo se reducirá. Los riñones desempeñan una función dominante en la regulación a largo plazo de la presión arterial al excretar cantidades variables de sodio y agua. Los riñones también contribuyen a la regulación a corto plazo de la presión arterial mediante la secreción de hormonas y factores o sustancias vasoactivos, como la renina.
Los riñones desempeñan una función dominante en la regulación a largo plazo de la presión arterial al excretar cantidades variables de sodio y agua. Los riñones también contribuyen a la regulación a corto plazo de la presión arterial mediante la secreción de hormonas y factores o sustancias vasoactivos, como la renina.
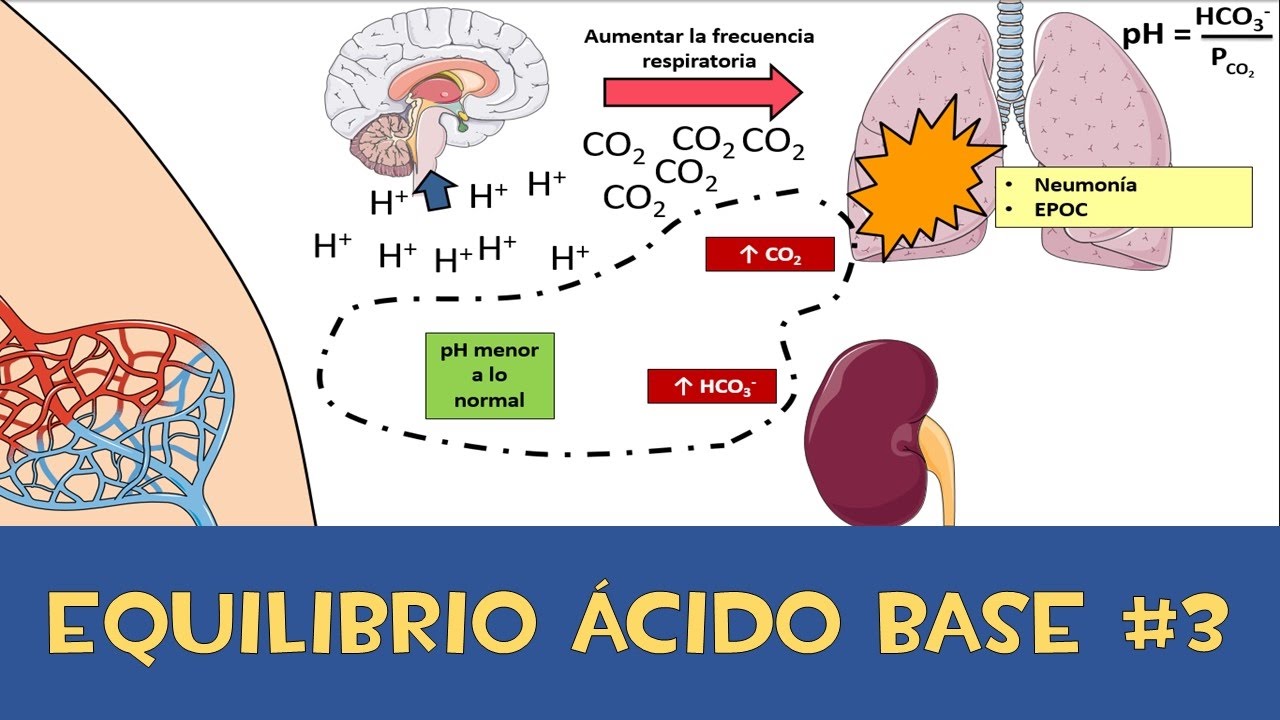

 Calcitriol es esencial para el depósito normal del calcio en el hueso y la reabsorción del calcio en el aparato digestivo.
Calcitriol es esencial para el depósito normal del calcio en el hueso y la reabsorción del calcio en el aparato digestivo.













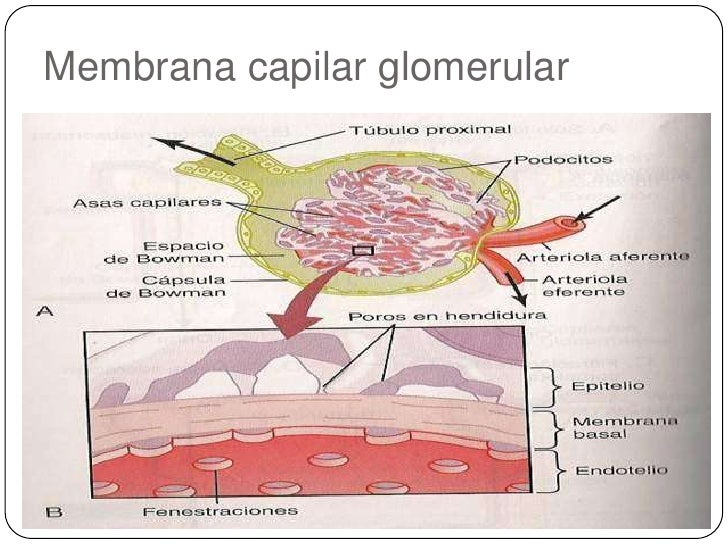 1) el endotelio del capilar. 2) una membrana basal. 3) una capa de células epiteliales (podocitos).
1) el endotelio del capilar. 2) una membrana basal. 3) una capa de células epiteliales (podocitos).



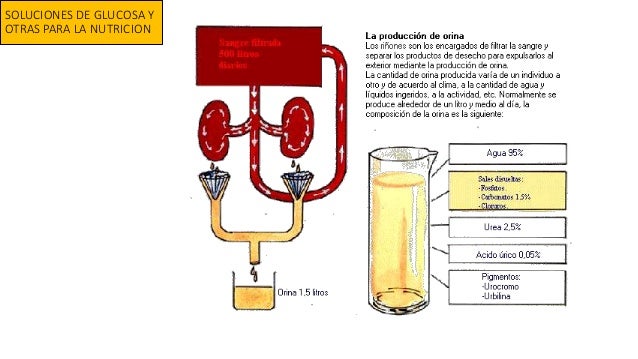
 Ingestión diaria de agua
Ingestión diaria de agua Pérdida insensible de agua: no se está consciente de esa perdida de agua, aunque se produce continuamente en todos los seres humanos vivos, por medio de las vías respiratorias y difusion a través de la piel.
Pérdida insensible de agua: no se está consciente de esa perdida de agua, aunque se produce continuamente en todos los seres humanos vivos, por medio de las vías respiratorias y difusion a través de la piel.


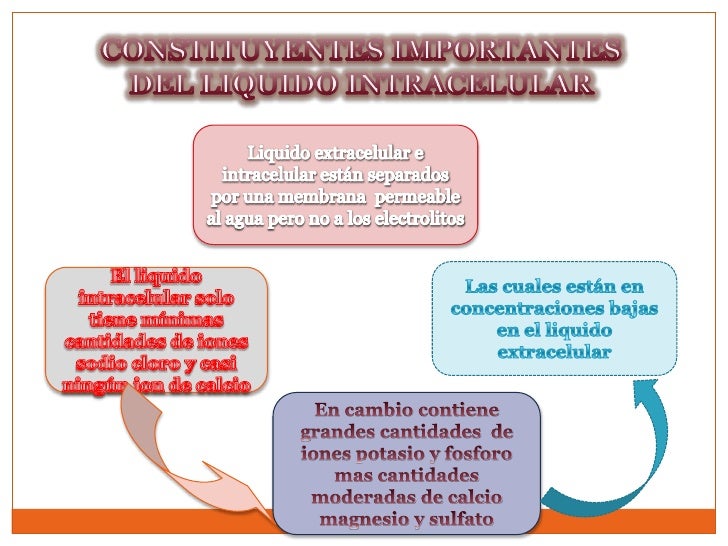
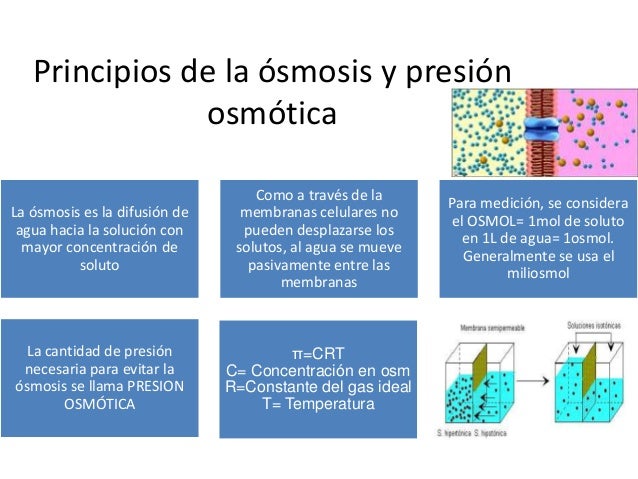
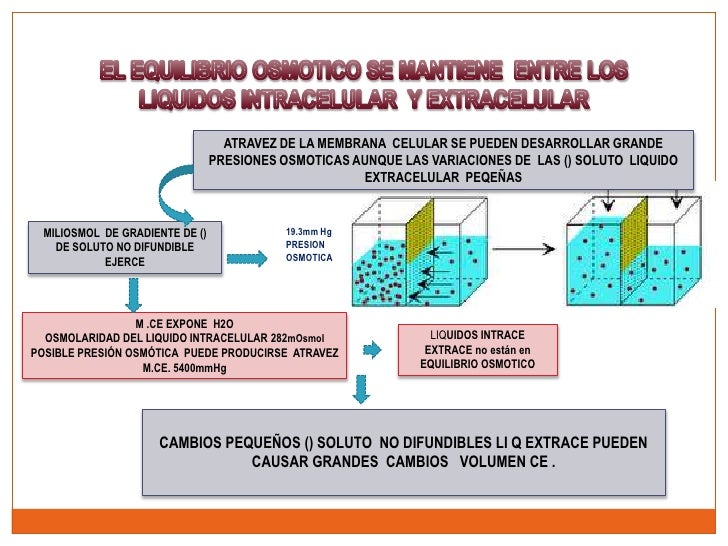
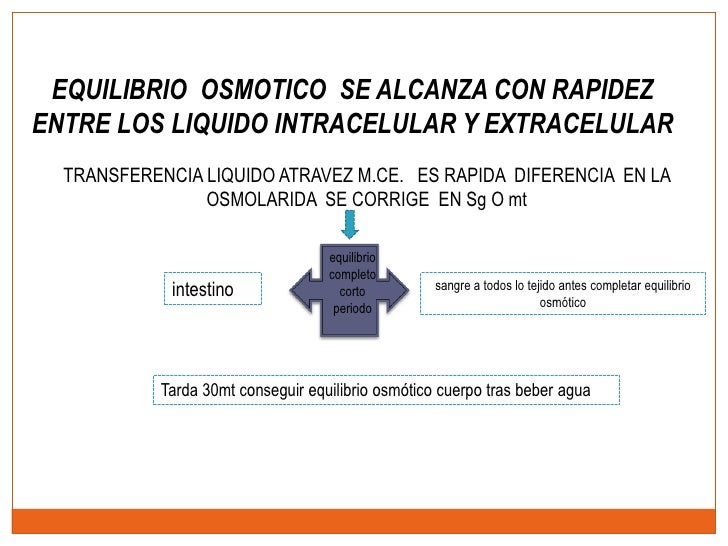
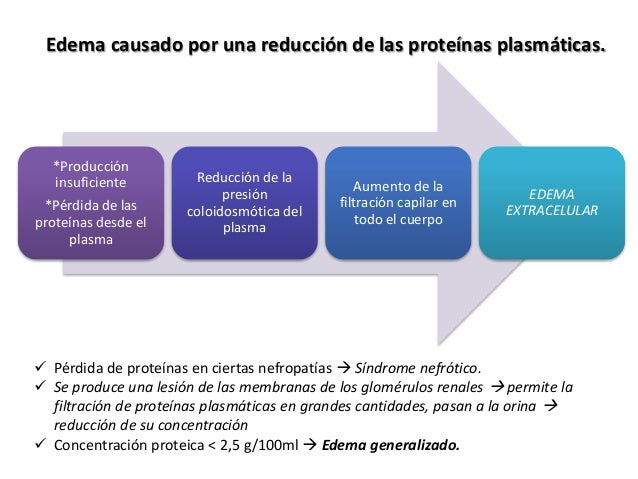
 El plasma es la parte no celular de la sangre; intercambia sustancias continuamente con el líquido intersticial a través de poros de las membranas capilares. Estos poros son muy permeables a casi todos los solutos del líquido extracelular excepto a las proteínas.
El plasma es la parte no celular de la sangre; intercambia sustancias continuamente con el líquido intersticial a través de poros de las membranas capilares. Estos poros son muy permeables a casi todos los solutos del líquido extracelular excepto a las proteínas.
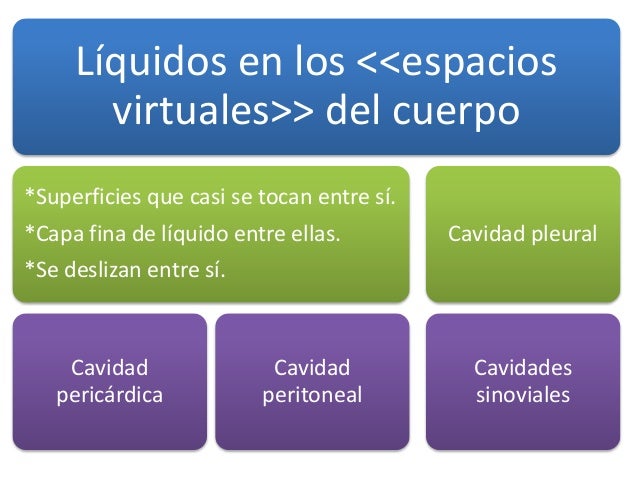
 La sangre contiene líquido extracelular (el líquido del plasma) y líquido intracelular (el líquido de los eritrocitos). Sin embargo, la sangre se considera un compartimiento líquido separado porque está contenida en su propia cámara, el aparato circulatorio. El volumen sanguíneo es especialmente importante en el control de la dinámica cardiovascular.
La sangre contiene líquido extracelular (el líquido del plasma) y líquido intracelular (el líquido de los eritrocitos). Sin embargo, la sangre se considera un compartimiento líquido separado porque está contenida en su propia cámara, el aparato circulatorio. El volumen sanguíneo es especialmente importante en el control de la dinámica cardiovascular.













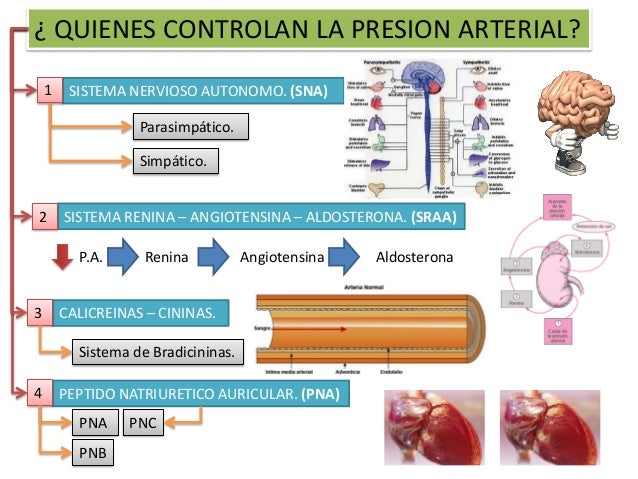
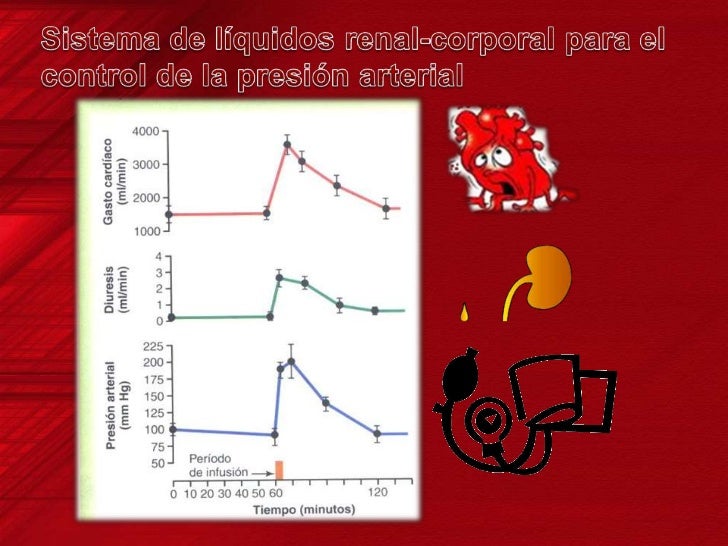
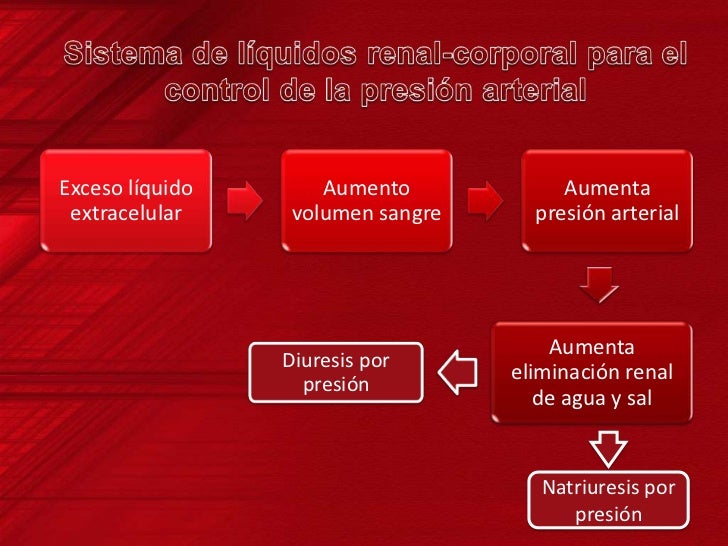

 Los dos determinantes principales de la presión arterial a largo plazo son los siguientes:
Los dos determinantes principales de la presión arterial a largo plazo son los siguientes: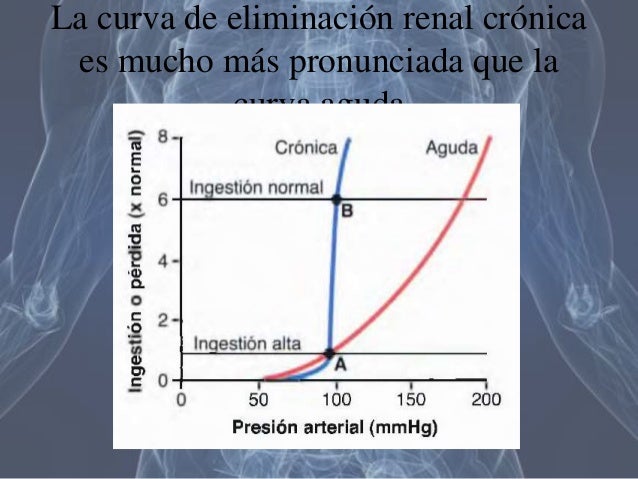









 La propia renina es una enzima y no una sustancia vasoactiva. La renina actúa enzimáticamente sobre otra proteína plasmática, una globulina denominada sustrato de renina (o angiotensinógeno), para liberar un péptido de 10 aminoácidos, la angio-tensina I, que tiene propiedades vasocons-trictoras discretas, no suficientes para provocar cambios suficientes en la función circulatoria. La renina persiste en la sangre durante 30 min hasta 1 h y continúa provocando la formación de aún más angiotensina I durante todo este tiempo.
La propia renina es una enzima y no una sustancia vasoactiva. La renina actúa enzimáticamente sobre otra proteína plasmática, una globulina denominada sustrato de renina (o angiotensinógeno), para liberar un péptido de 10 aminoácidos, la angio-tensina I, que tiene propiedades vasocons-trictoras discretas, no suficientes para provocar cambios suficientes en la función circulatoria. La renina persiste en la sangre durante 30 min hasta 1 h y continúa provocando la formación de aún más angiotensina I durante todo este tiempo.





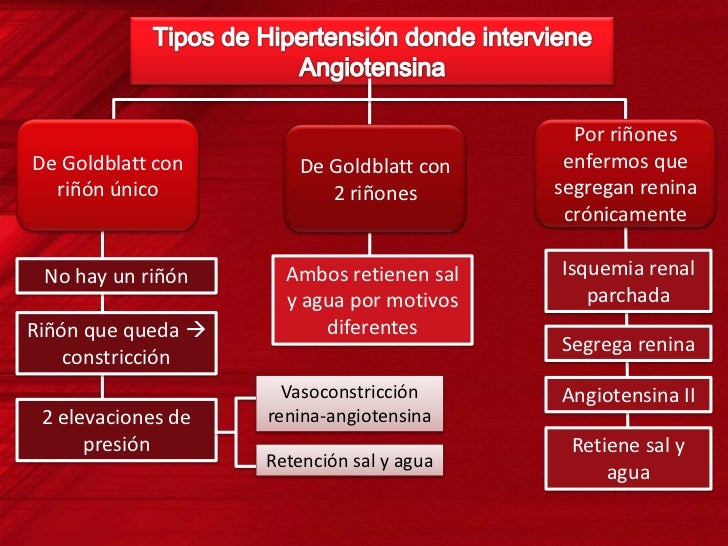

 El
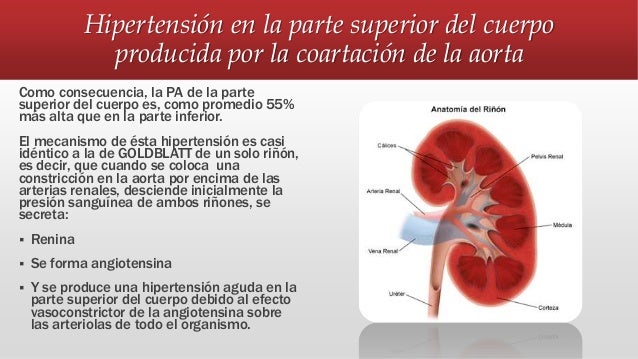
El 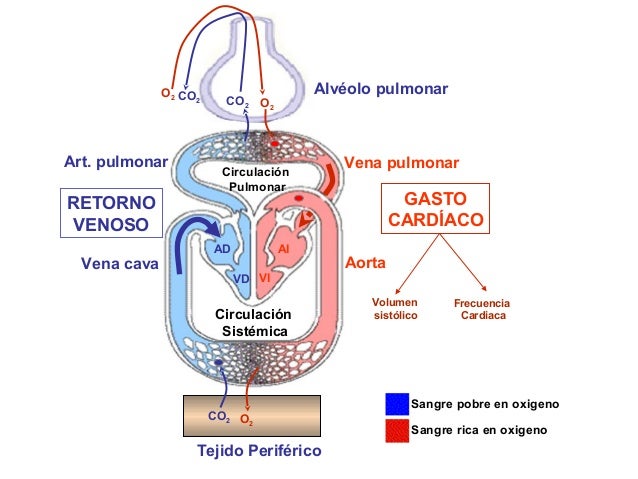
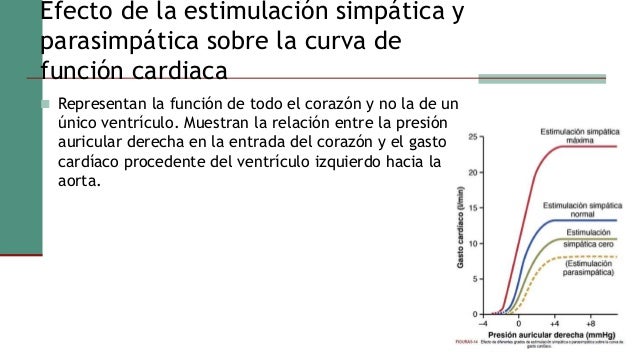
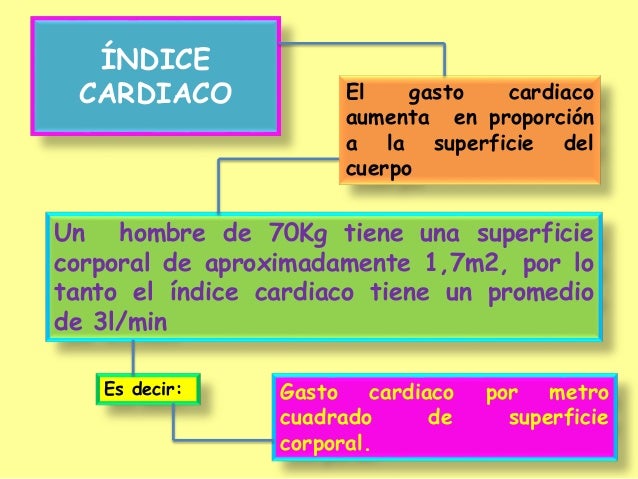 En estudios experimentales se ha demostrado que el gasto cardíaco aumenta en proporción a la superficie corporal. el gasto cardíaco se expresa en términos de índice cardíaco, que es el gasto cardíaco por metro cuadrado de superficie corporal.
En estudios experimentales se ha demostrado que el gasto cardíaco aumenta en proporción a la superficie corporal. el gasto cardíaco se expresa en términos de índice cardíaco, que es el gasto cardíaco por metro cuadrado de superficie corporal.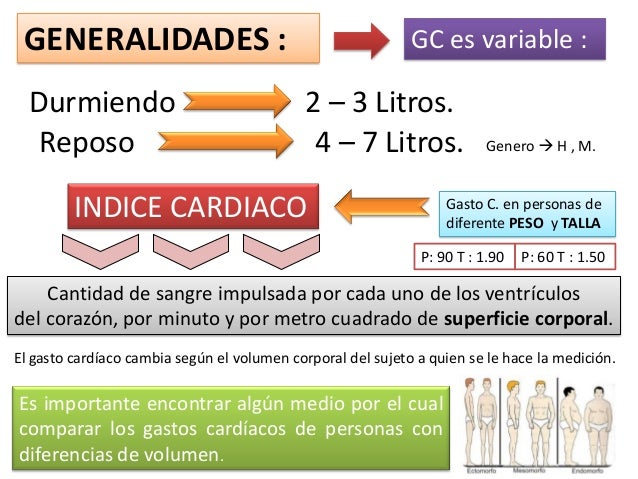

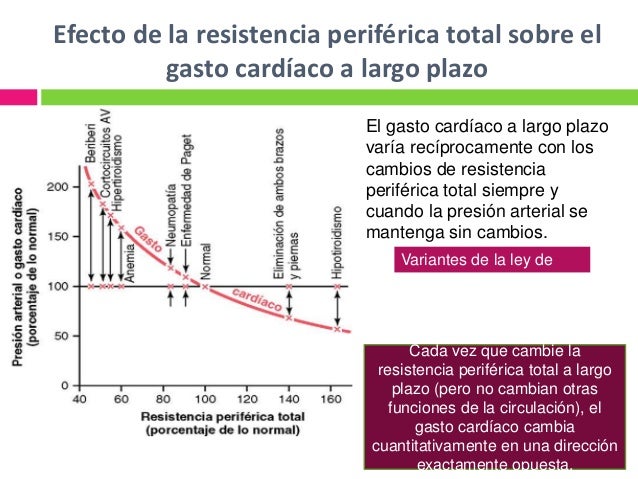
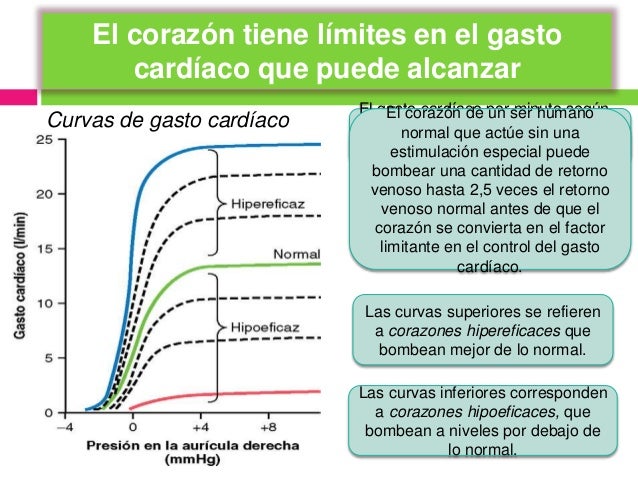


 Durante el ejercicio, el aumento intenso del metabolismo en los músculos esqueléticos activos actúa directamente en las arteriolas musculares para relajarlos y permitir el acceso adecuado del oxígeno y otros nutrientes necesarios para mantener la contracción muscular. Así se produce un descenso importante de la resistencia periférica total, lo que normalmente también disminuiría la presión arterial.
Durante el ejercicio, el aumento intenso del metabolismo en los músculos esqueléticos activos actúa directamente en las arteriolas musculares para relajarlos y permitir el acceso adecuado del oxígeno y otros nutrientes necesarios para mantener la contracción muscular. Así se produce un descenso importante de la resistencia periférica total, lo que normalmente también disminuiría la presión arterial.

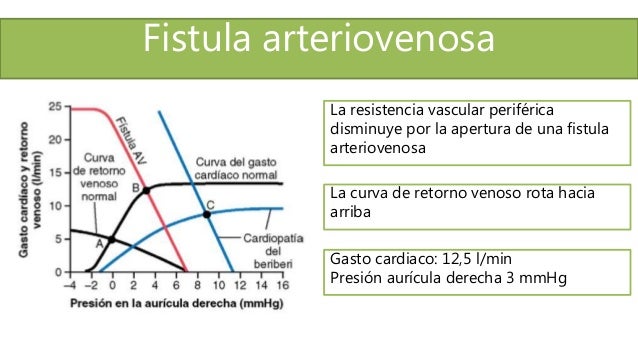
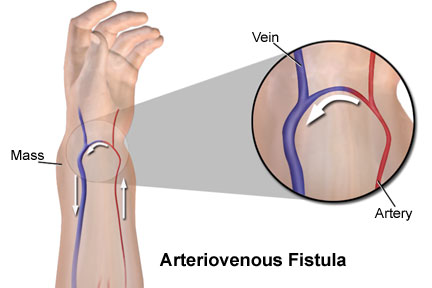 Cada vez que se crea una fístula (cortocircuito AV) entre una arteria y una vena importantes pasa una cantidad enorme de flujo sanguíneo directamente desde la arteria hacia la vena, lo cual, disminuye en gran medida la resistencia periférica total y, así mismo, aumenta el retorno venoso y el gasto cardíaco.
Cada vez que se crea una fístula (cortocircuito AV) entre una arteria y una vena importantes pasa una cantidad enorme de flujo sanguíneo directamente desde la arteria hacia la vena, lo cual, disminuye en gran medida la resistencia periférica total y, así mismo, aumenta el retorno venoso y el gasto cardíaco. En el hipertiroidismo, el metabolismo de la mayoría de los tejidos del organismo está muy aumentado y la utilización de oxígeno aumenta, liberándose productos vasodila-tadores desde los tejidos. Por tanto, la resistencia periférica total disminuye mucho.
En el hipertiroidismo, el metabolismo de la mayoría de los tejidos del organismo está muy aumentado y la utilización de oxígeno aumenta, liberándose productos vasodila-tadores desde los tejidos. Por tanto, la resistencia periférica total disminuye mucho.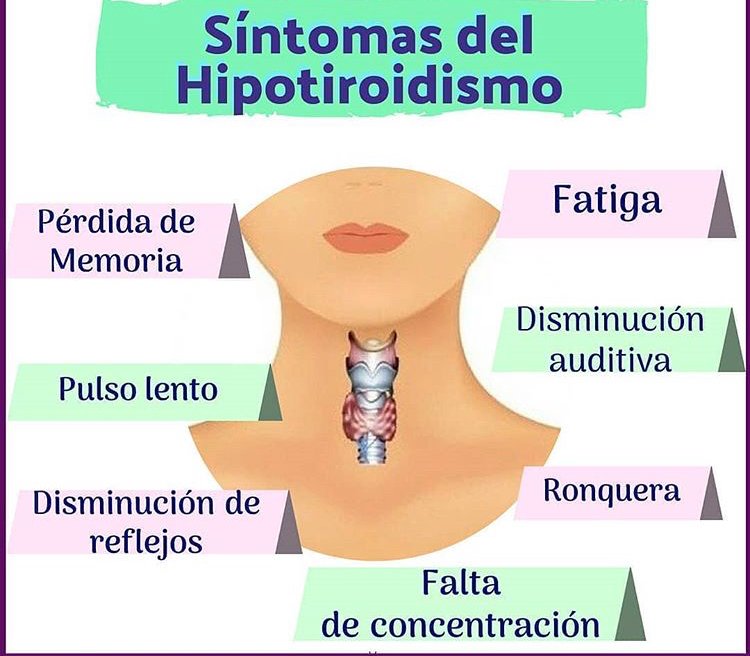
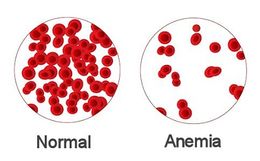 Se producen dos efectos periféricos que disminuyen en gran medida la resistencia periférica total.
Se producen dos efectos periféricos que disminuyen en gran medida la resistencia periférica total.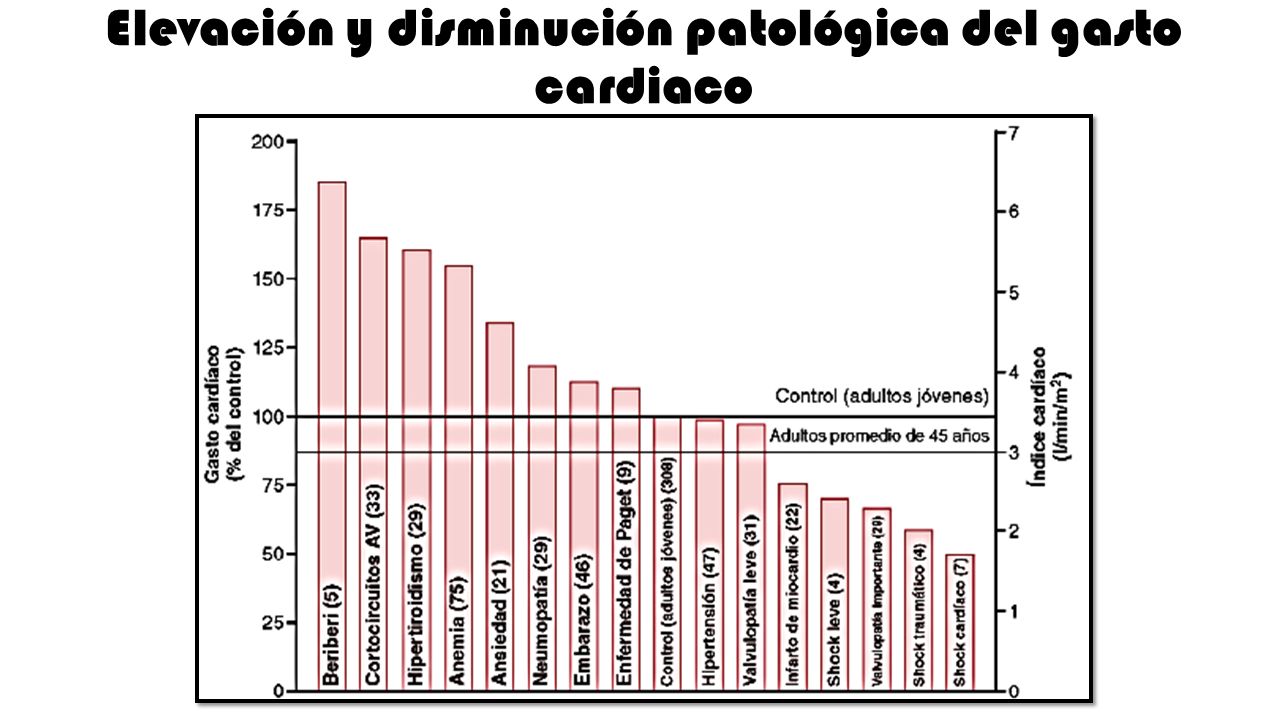
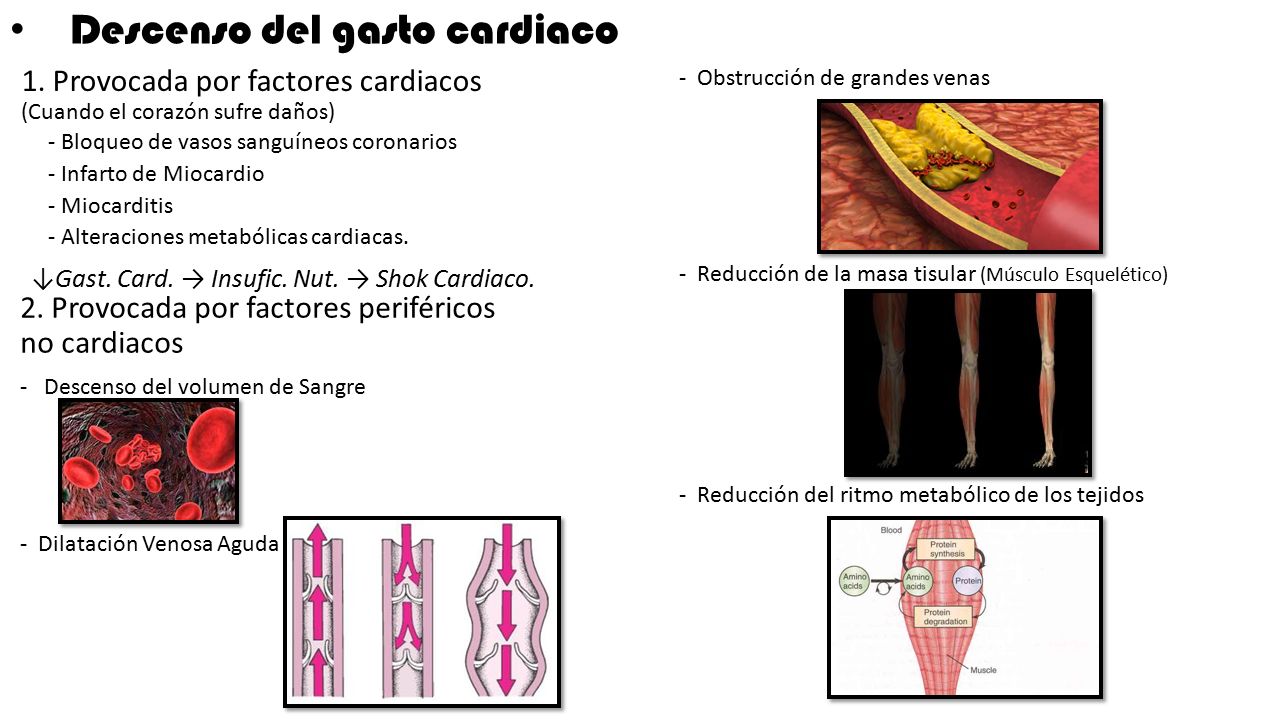

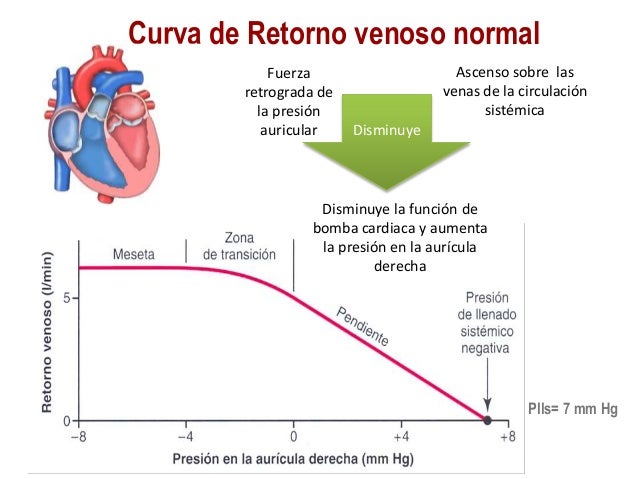
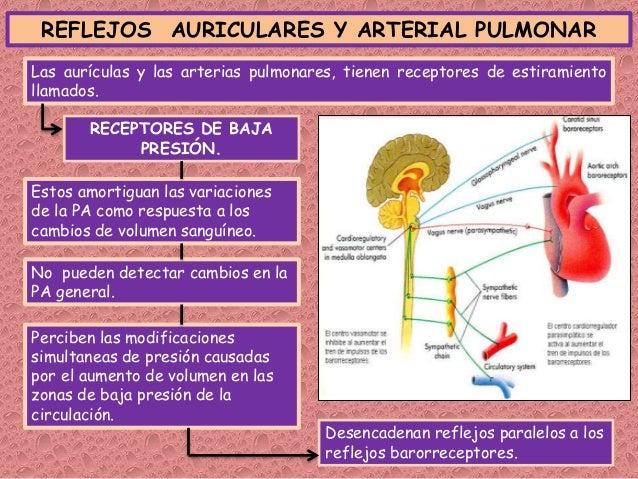
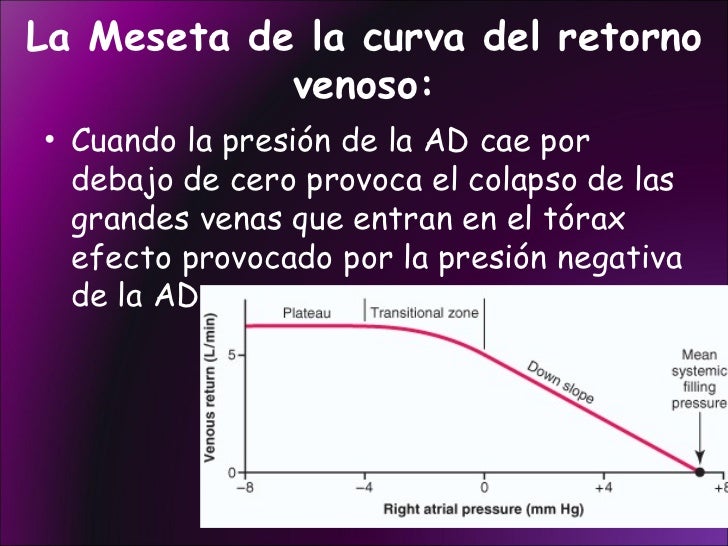 Cuando la presión en la aurícula derecha cae por debajo de cero, es decir, por debajo de la presión atmosférica, aumenta más cuando cesa casi totalmente el retorno venoso. Y el retorno venoso habrá alcanzado la meseta en el momento en el que la presión en la aurícula derecha haya caído hasta –2 mm y se mantiene en la meseta aunque la presión en la aurícula derecha caiga hasta –20 mmHg, –50 mmHg o incluso más. Esta meseta está provocada por el colapso de las venas que entran en el tórax.
Cuando la presión en la aurícula derecha cae por debajo de cero, es decir, por debajo de la presión atmosférica, aumenta más cuando cesa casi totalmente el retorno venoso. Y el retorno venoso habrá alcanzado la meseta en el momento en el que la presión en la aurícula derecha haya caído hasta –2 mm y se mantiene en la meseta aunque la presión en la aurícula derecha caiga hasta –20 mmHg, –50 mmHg o incluso más. Esta meseta está provocada por el colapso de las venas que entran en el tórax.
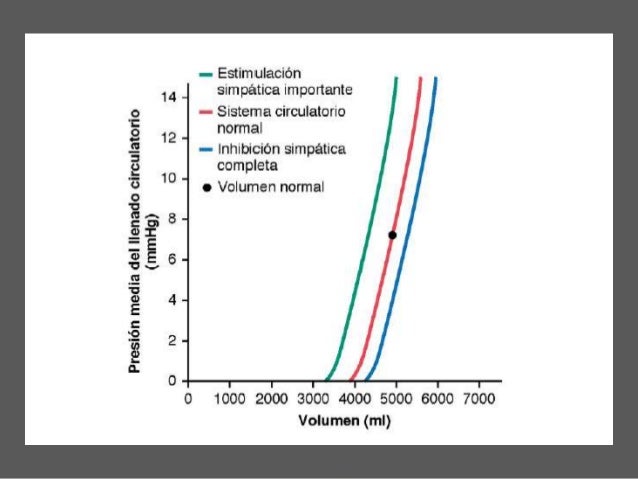
 La presión media del llenado sistémico (Plls) es algo diferente de la presión media del llenado circulatorio, ya que es la presión media en cualquier punto de la circulación sistémica después de que el flujo sanguíneo se haya interrumpido al pinzar los vasos sanguíneos grandes en el corazón, por lo que se puede medir la presión de la circulación sistémica independientemente de la presión que haya en la circulación pulmonar.
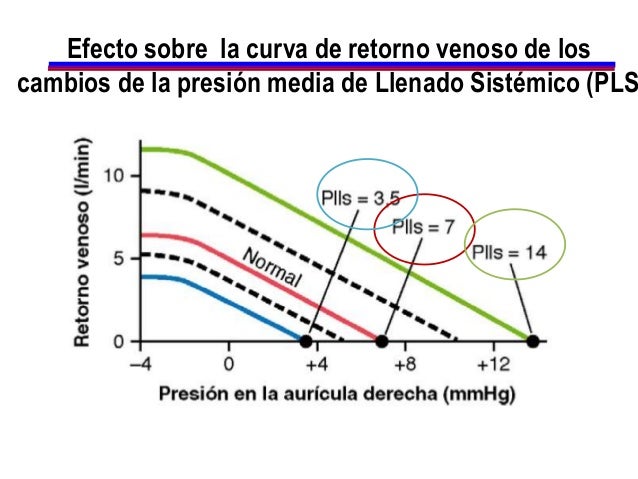
La presión media del llenado sistémico (Plls) es algo diferente de la presión media del llenado circulatorio, ya que es la presión media en cualquier punto de la circulación sistémica después de que el flujo sanguíneo se haya interrumpido al pinzar los vasos sanguíneos grandes en el corazón, por lo que se puede medir la presión de la circulación sistémica independientemente de la presión que haya en la circulación pulmonar.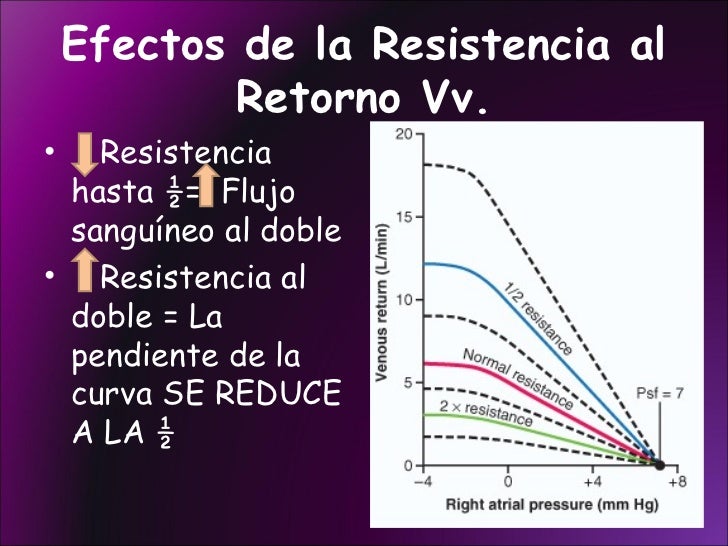
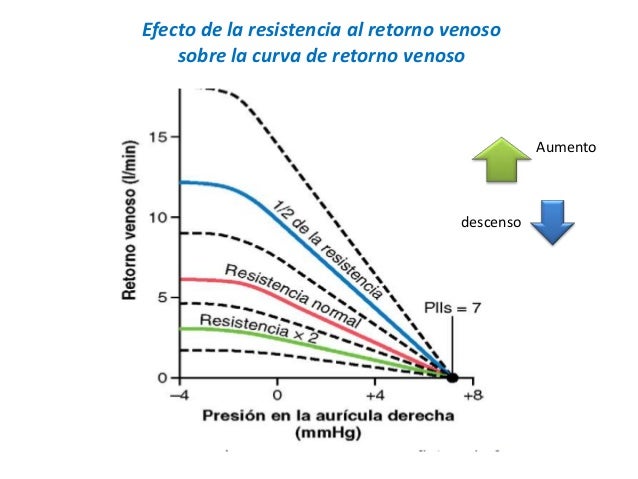
 Cuando actúa la circulación completa, el corazón y la circulación sistémica deben funcionar conjuntamente, lo que significa que:
Cuando actúa la circulación completa, el corazón y la circulación sistémica deben funcionar conjuntamente, lo que significa que:





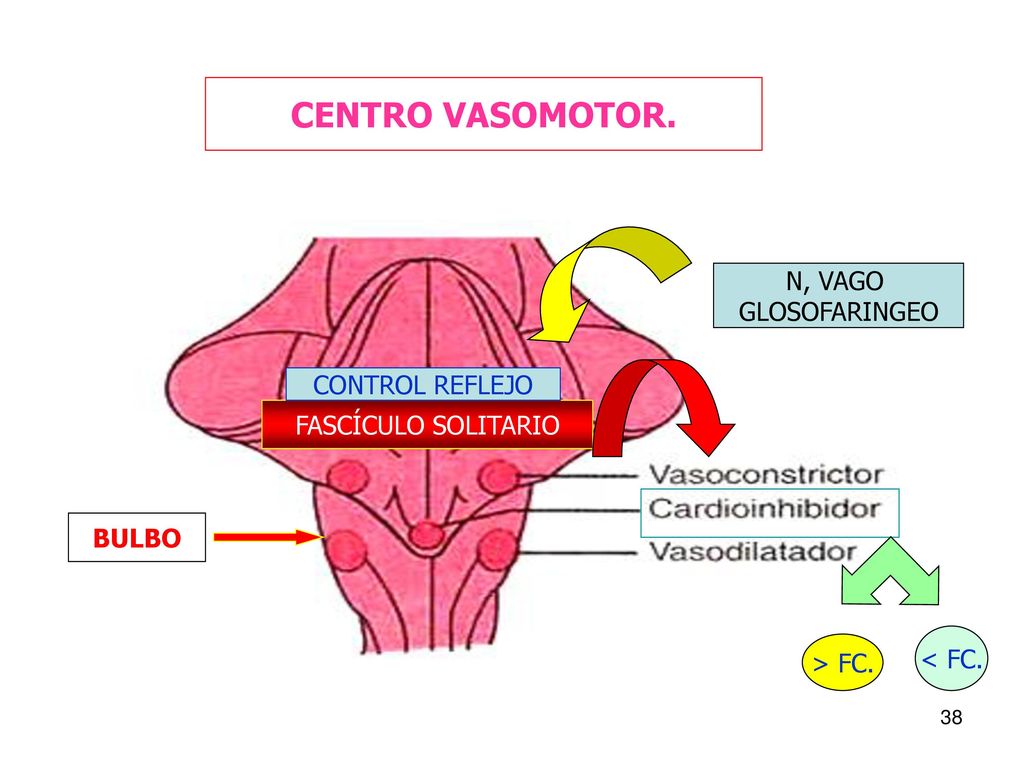


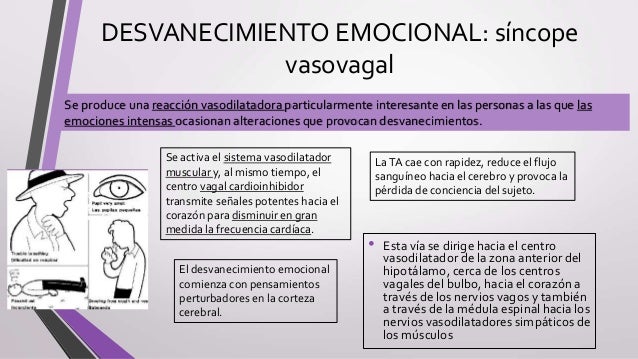
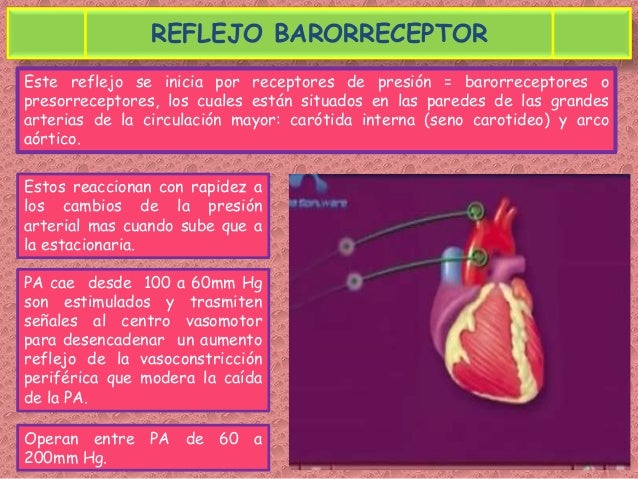
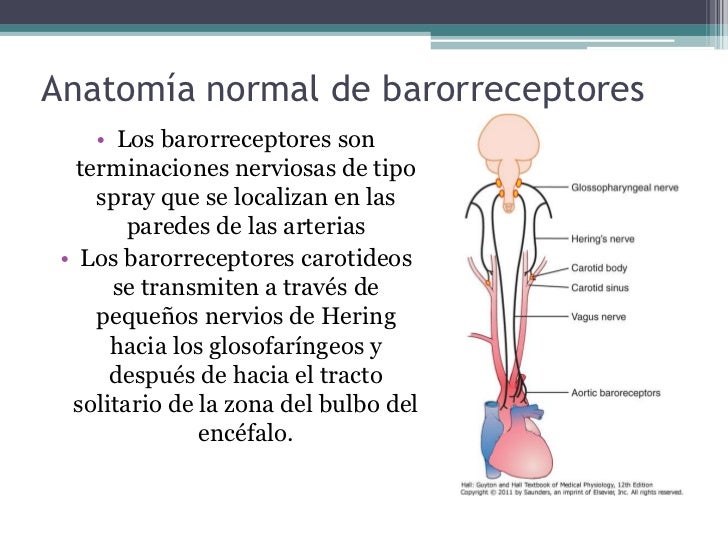
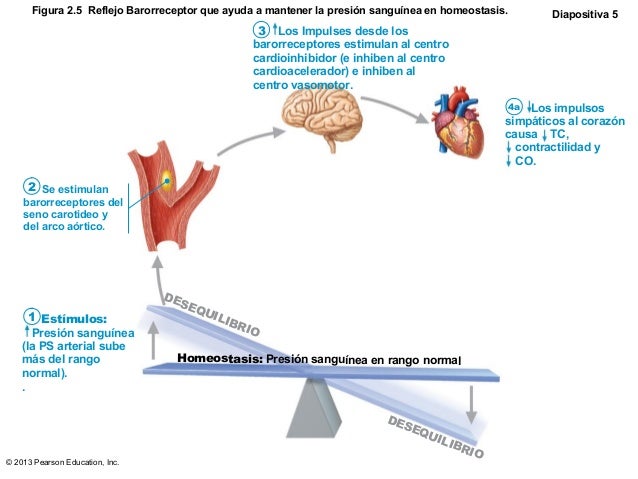





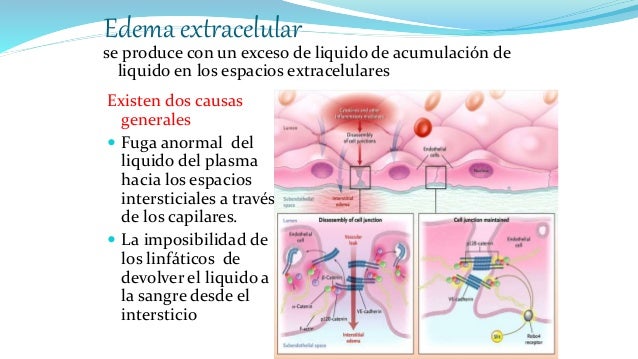
 El sistema linfático representa una vía accesoria a través de la cual el líquido puede fluir desde los espacios intersticiales hacia la sangre. El sistema linfático también es una de las vías principales de absorción de los nutrientes del aparato digestivo, en especial de la absorción de prácticamente todas las grasas del alimento. Los vasos linfáticos transportan las proteínas y las macropartículas de los espacios tisulares, ya que ninguna de las cuales puede ser eliminada por absorción directamente hacia los capilares sanguíneos.
El sistema linfático representa una vía accesoria a través de la cual el líquido puede fluir desde los espacios intersticiales hacia la sangre. El sistema linfático también es una de las vías principales de absorción de los nutrientes del aparato digestivo, en especial de la absorción de prácticamente todas las grasas del alimento. Los vasos linfáticos transportan las proteínas y las macropartículas de los espacios tisulares, ya que ninguna de las cuales puede ser eliminada por absorción directamente hacia los capilares sanguíneos. La mayoría de los tejidos del prgnaismo tiene vasos linfáticos, sinmebargo, hay algunas excepciones, como las porciones superficiales de la piel, el sistema nervioso central y el endomisio de músculos y huesos. Pero incluso estos tejidos tienen canales intersticiales diminutos que se denominan canales prelinfáticos.
La mayoría de los tejidos del prgnaismo tiene vasos linfáticos, sinmebargo, hay algunas excepciones, como las porciones superficiales de la piel, el sistema nervioso central y el endomisio de músculos y huesos. Pero incluso estos tejidos tienen canales intersticiales diminutos que se denominan canales prelinfáticos.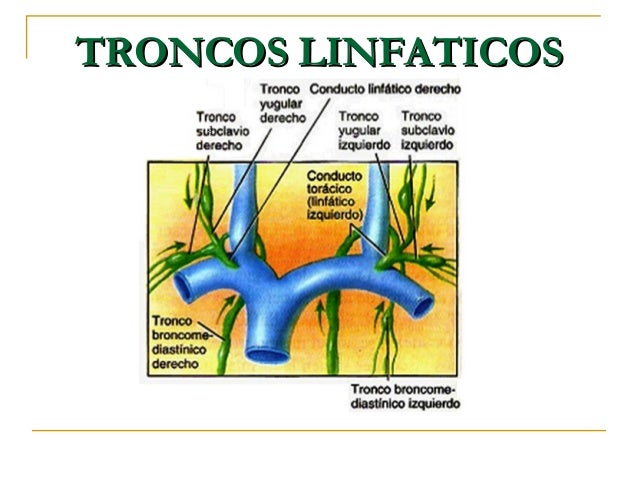 Todos los vasos linfáticos de la mitad inferior del organismo se vaciarán en el conducto torácico, que a su vez se vacía en el sistema venoso en la unión de la vena yugular interna con la vena subclavia izquierda.
Todos los vasos linfáticos de la mitad inferior del organismo se vaciarán en el conducto torácico, que a su vez se vacía en el sistema venoso en la unión de la vena yugular interna con la vena subclavia izquierda.
 La linfa deriva del líquido intersticial que fluye en los linfáticos. La concentración de proteínas en el líquido intersticial de la mayoría de los tejidos alcanza un promedio de 2 g/dl y la concentración de proteínas del flujo linfático que procede de
La linfa deriva del líquido intersticial que fluye en los linfáticos. La concentración de proteínas en el líquido intersticial de la mayoría de los tejidos alcanza un promedio de 2 g/dl y la concentración de proteínas del flujo linfático que procede de

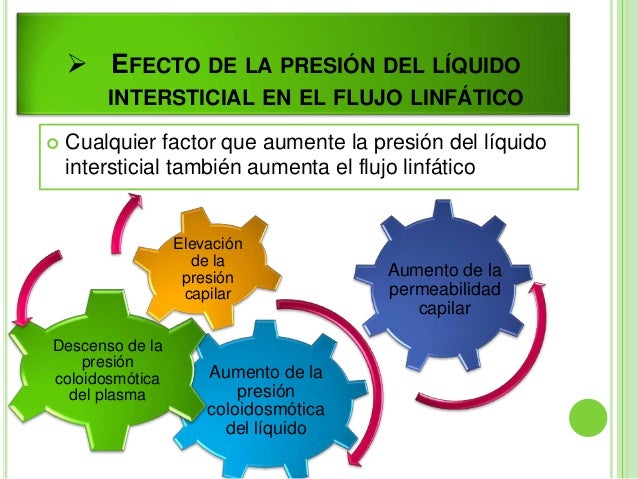

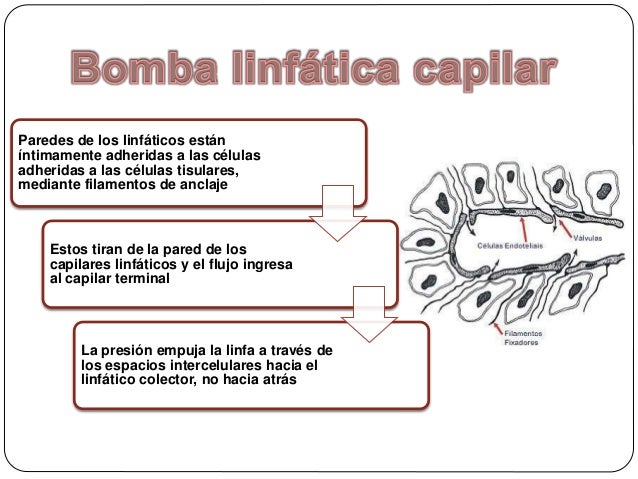


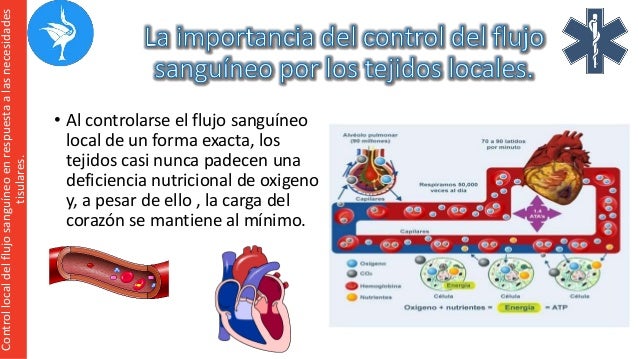



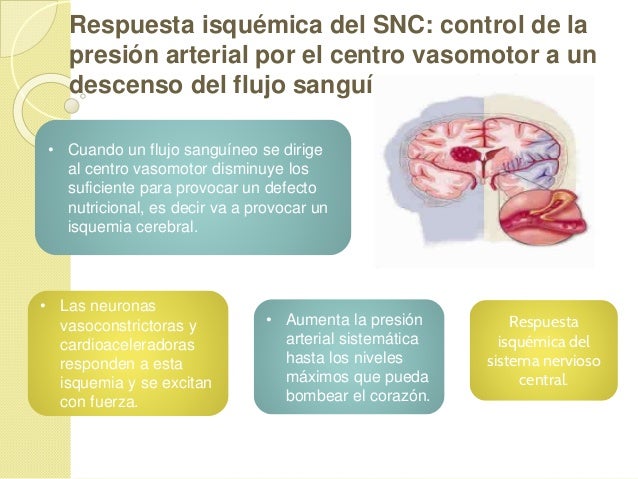
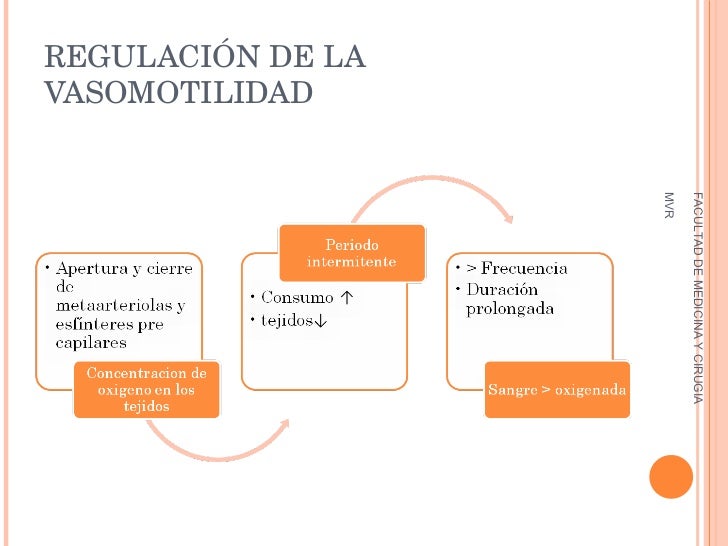
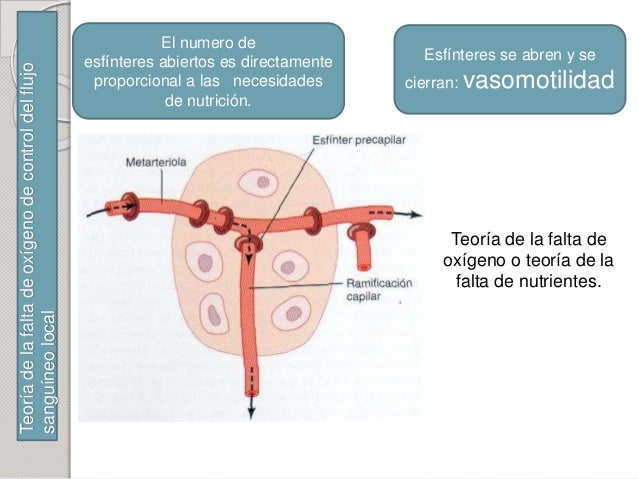
 Establece que cuanto mayor sea el metabo-lismo o menor sea la disponibilidad de oxígeno o de algunos otros nutrien tes en un tejido, mayor será la velocidad de formación de sustancias vasodila-tadoras en las células de ese tejido. Se cree que estas sustancias vasodilatadoras di-funden a través de los tejidos hacia los esfínteres precapi-lares, las metaarteriolas y las arteriolas para provocar la dilatación. Se han propuesto varias sustancias vasodilatadoras diferentes, como adenosina, dióxido de carbono, compuestos con fosfato de adenosina, histamina, iones potasio e iones hidrógeno.
Establece que cuanto mayor sea el metabo-lismo o menor sea la disponibilidad de oxígeno o de algunos otros nutrien tes en un tejido, mayor será la velocidad de formación de sustancias vasodila-tadoras en las células de ese tejido. Se cree que estas sustancias vasodilatadoras di-funden a través de los tejidos hacia los esfínteres precapi-lares, las metaarteriolas y las arteriolas para provocar la dilatación. Se han propuesto varias sustancias vasodilatadoras diferentes, como adenosina, dióxido de carbono, compuestos con fosfato de adenosina, histamina, iones potasio e iones hidrógeno.


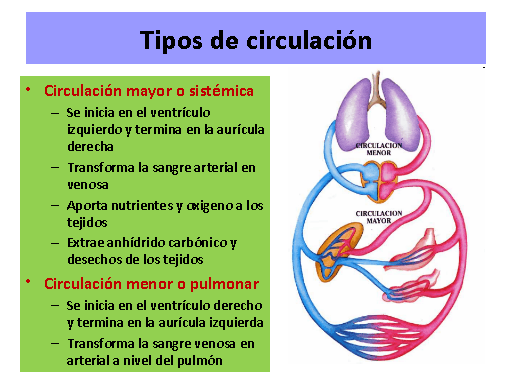 Está divida en circulación sistémica y circulación pulmonar. Como la circulación sisté-mica aporta el flujo sanguíneo a todos los tejidos del organismo excepto los pulmones, también se conoce
Está divida en circulación sistémica y circulación pulmonar. Como la circulación sisté-mica aporta el flujo sanguíneo a todos los tejidos del organismo excepto los pulmones, también se conoce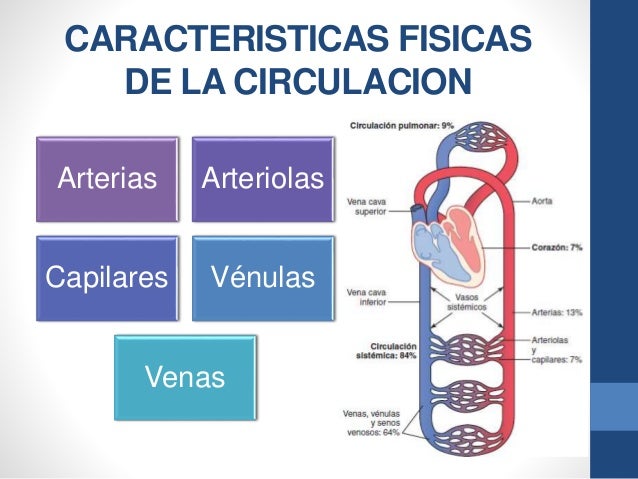


 El corazón bombea la sangre continuamente hacia la aorta, la presión media en este vaso es alta, con una media en torno a los 100 mmHg. Además, como el bombeo cardíaco es pulsátil, la presión arterial alterna entre una presión sistólica de 120 mmHg y una diastólica de 80 mmHg. A medida que el flujo sanguíneo atraviesa la circulación sistémica la presión media va cayendo progresivamente hasta llegar casi a 0 mmHg en el momento en el que alcanza la terminación de las venas cava, donde se vacía en la aurícula derecha del corazón.
El corazón bombea la sangre continuamente hacia la aorta, la presión media en este vaso es alta, con una media en torno a los 100 mmHg. Además, como el bombeo cardíaco es pulsátil, la presión arterial alterna entre una presión sistólica de 120 mmHg y una diastólica de 80 mmHg. A medida que el flujo sanguíneo atraviesa la circulación sistémica la presión media va cayendo progresivamente hasta llegar casi a 0 mmHg en el momento en el que alcanza la terminación de las venas cava, donde se vacía en la aurícula derecha del corazón.

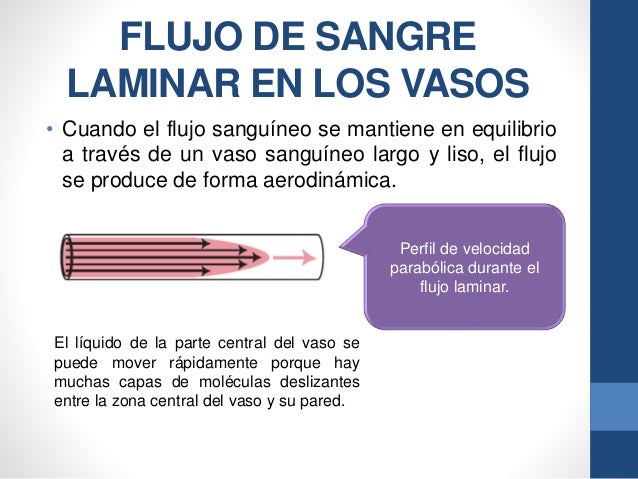


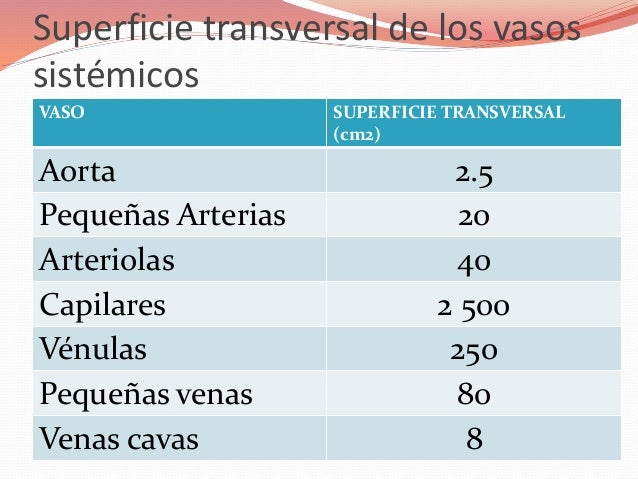
 La velocidad del flujo sanguíneo a través de todo el sistema circulatorio es igual a la velocidad de la sangre que bombea el corazón, es decir, es igual al gasto cardíaco. En un ser humano adulto es aproximadamente igual a 100 ml/s.
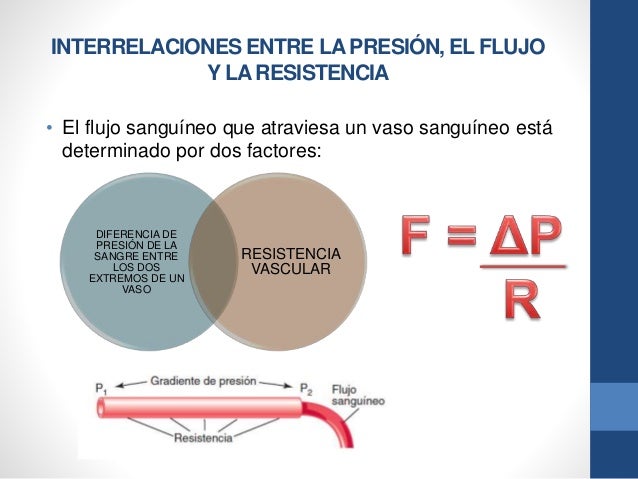
La velocidad del flujo sanguíneo a través de todo el sistema circulatorio es igual a la velocidad de la sangre que bombea el corazón, es decir, es igual al gasto cardíaco. En un ser humano adulto es aproximadamente igual a 100 ml/s. La sangre que bombea el corazón fluye desde la parte de presión alta de la circulación sistémica (la aorta) hacia el lado de baja presión (la vena cava) a través de muchos miles de vasos sanguíneos dispuestos en serie y en paralelo. Las arterias, arteriolas, capilares, vénulas y venas se disponen colectivamente en serie. Es decir, la resistencia vascular periférica total es igual a la suma de resistencias de las arterias, arteriolas, capilares, vénulas y venas.
La sangre que bombea el corazón fluye desde la parte de presión alta de la circulación sistémica (la aorta) hacia el lado de baja presión (la vena cava) a través de muchos miles de vasos sanguíneos dispuestos en serie y en paralelo. Las arterias, arteriolas, capilares, vénulas y venas se disponen colectivamente en serie. Es decir, la resistencia vascular periférica total es igual a la suma de resistencias de las arterias, arteriolas, capilares, vénulas y venas.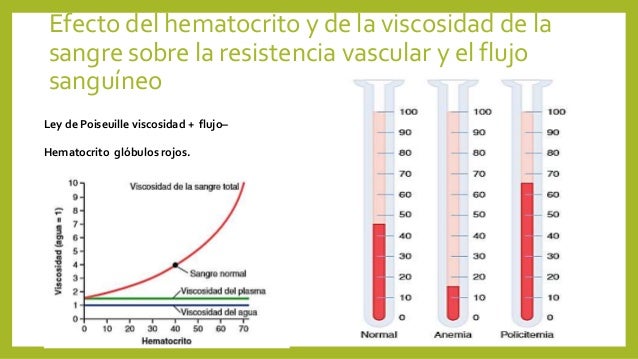 La viscosidad de la sangre aumenta drásti-camente a medida que lo hace el hematocrito. La viscosidad de la sangre total con un hematocrito normal es de 3.
La viscosidad de la sangre aumenta drásti-camente a medida que lo hace el hematocrito. La viscosidad de la sangre total con un hematocrito normal es de 3.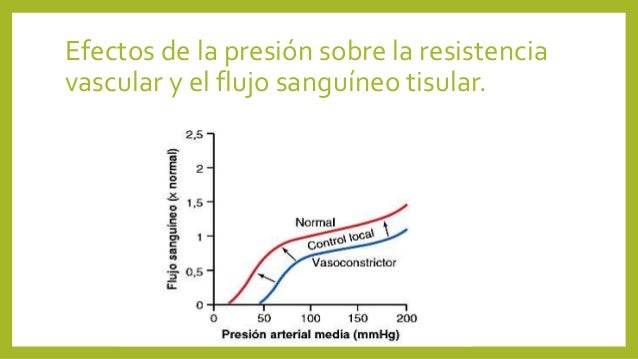
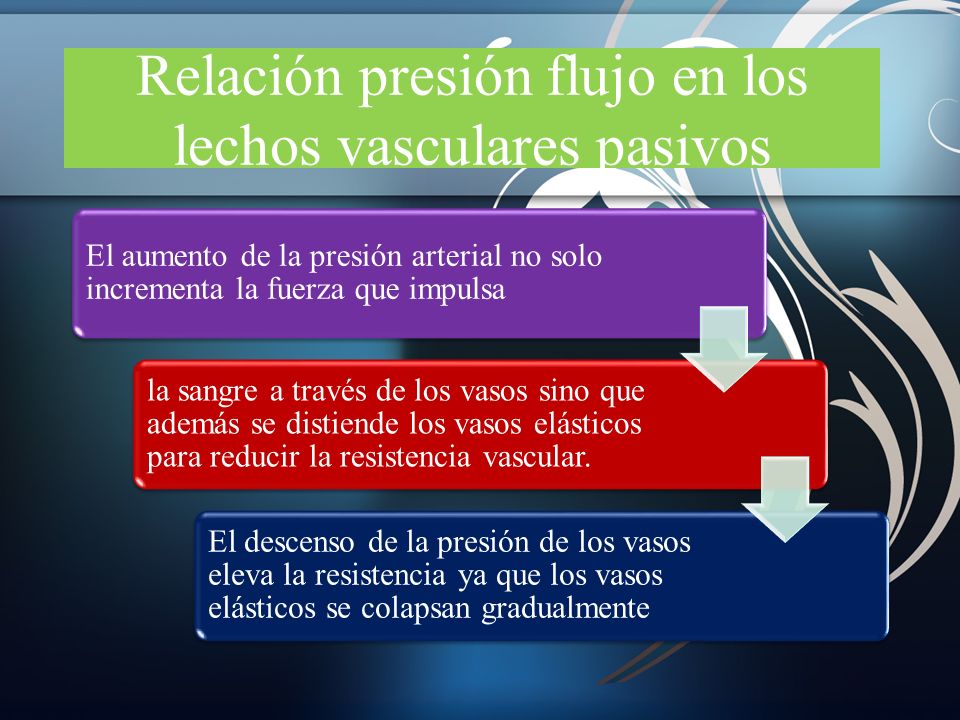
 El nódulo sinusal (nódulo sinoauricular) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Las fibras de este nódulo casi no tienen filamentos musculares contráctiles y cada una de ellas tiene sólo de 3 a 5 mm de diámetro, en contraste con un diámetro de 10 a 15 mm para las fibras musculares auriculares circundantes. Las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.
El nódulo sinusal (nódulo sinoauricular) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Las fibras de este nódulo casi no tienen filamentos musculares contráctiles y cada una de ellas tiene sólo de 3 a 5 mm de diámetro, en contraste con un diámetro de 10 a 15 mm para las fibras musculares auriculares circundantes. Las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.

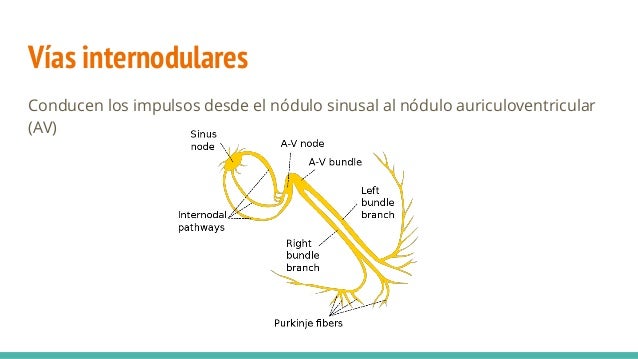 Los extremos de las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares circundantes, por tanto, los potenciales de acción que se originan en el nódulo sinusal viajan hacia estas fibras musculares auriculares,así el potencial de acción se propaga por toda la masa muscular auricular y, finalmente, llega hasta el nódulo AV. La velocidad de conducción en la mayor parte del músculo auricular es de aproximadamente 0,3 m/s, pero la conducción es más rápida, de aproximadamente 1 m/s, en varias pequeñas bandas de fibras auriculares, ejemplos de ello son las vías internodulares anterior, media y posterior. La causa de la velocidad de conducción más rápida de estas bandas es
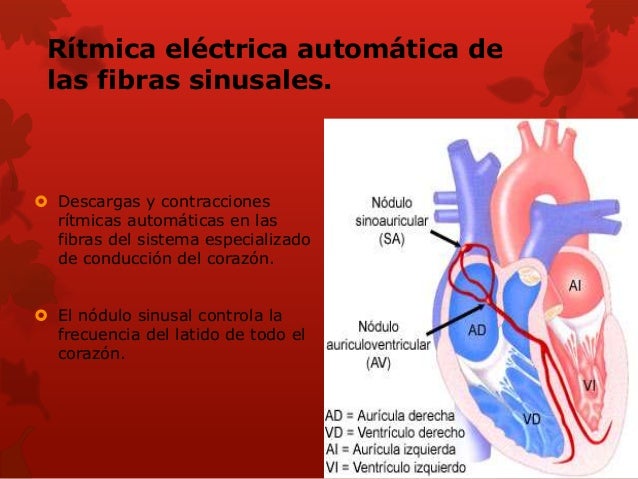
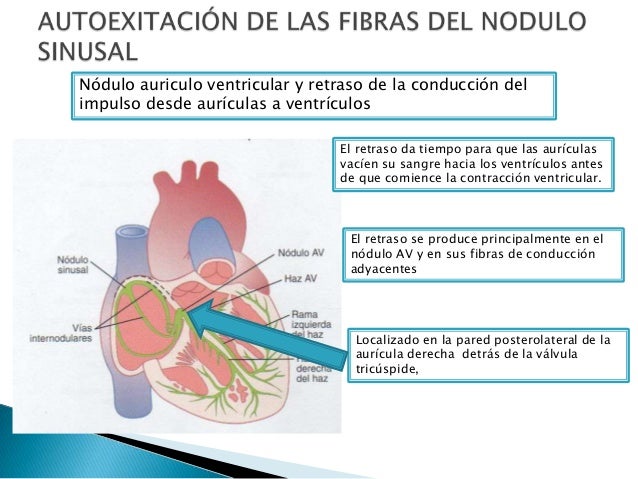
Los extremos de las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares circundantes, por tanto, los potenciales de acción que se originan en el nódulo sinusal viajan hacia estas fibras musculares auriculares,así el potencial de acción se propaga por toda la masa muscular auricular y, finalmente, llega hasta el nódulo AV. La velocidad de conducción en la mayor parte del músculo auricular es de aproximadamente 0,3 m/s, pero la conducción es más rápida, de aproximadamente 1 m/s, en varias pequeñas bandas de fibras auriculares, ejemplos de ello son las vías internodulares anterior, media y posterior. La causa de la velocidad de conducción más rápida de estas bandas es El sistema de conducción auricular está organizado de modo que el impulso cardíaco no viaja desde las aurículas hacia los ventrículos demasiado rápidamente; este retraso da tiempo para que las aurículas vacíen su sangre hacia los ventrículos antes de que comience la contracción ventricular. El retraso de la transmisión hacia los ventrículos se produce principalmente en el nódulo AV y en sus fibras de conducción adyacentes.
El sistema de conducción auricular está organizado de modo que el impulso cardíaco no viaja desde las aurículas hacia los ventrículos demasiado rápidamente; este retraso da tiempo para que las aurículas vacíen su sangre hacia los ventrículos antes de que comience la contracción ventricular. El retraso de la transmisión hacia los ventrículos se produce principalmente en el nódulo AV y en sus fibras de conducción adyacentes.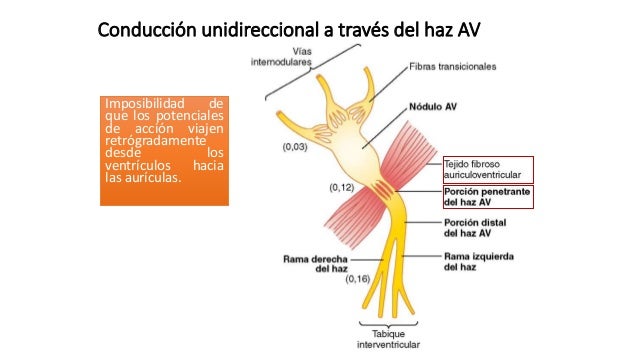 Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógra-damente desde los ventrículos hacia las aurículas. Esto impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo sólo la contracción anterógrada desde las aurículas hacia los ventrículos; el haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa continua, ésta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV.
Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógra-damente desde los ventrículos hacia las aurículas. Esto impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo sólo la contracción anterógrada desde las aurículas hacia los ventrículos; el haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa continua, ésta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV. Los extremos de las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas. Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de sólo 0,03 s.
Los extremos de las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas. Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de sólo 0,03 s.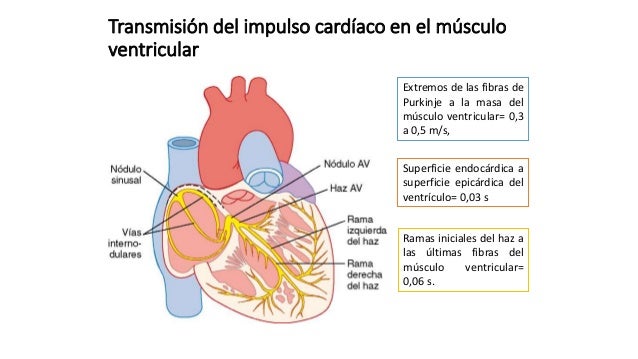 Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora sólo de 0,3 a 0,5 m/s.
Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora sólo de 0,3 a 0,5 m/s.
 El impulso normalmente se origina en el nódulo sinusal, por lo que es el que controla la ritmicidad del corazón. Esto se debe a que la frecuencia de descarga del nódulo sinusal es considerablemente mayor que la frecuencia de descarga autoexcitadora natural de las fibras del nódulo AV y de las fibras de Purkinje. Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo AV y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. Pero el nódulo sinusal produce una nueva descarga antes de que las fibras del nódulo AV o las fibras de Purkinje puedan alcanzar sus propios umbrales de autoexcitación. Así, el nódulo sinusal controla el latido del corazón porque su frecuencia de descarga rítmica es más rápida que la de cualquier otra parte del corazón. Por tanto, el nódulo sinusal es prácticamente siempre el marcapasos del corazón normal.
El impulso normalmente se origina en el nódulo sinusal, por lo que es el que controla la ritmicidad del corazón. Esto se debe a que la frecuencia de descarga del nódulo sinusal es considerablemente mayor que la frecuencia de descarga autoexcitadora natural de las fibras del nódulo AV y de las fibras de Purkinje. Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo AV y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. Pero el nódulo sinusal produce una nueva descarga antes de que las fibras del nódulo AV o las fibras de Purkinje puedan alcanzar sus propios umbrales de autoexcitación. Así, el nódulo sinusal controla el latido del corazón porque su frecuencia de descarga rítmica es más rápida que la de cualquier otra parte del corazón. Por tanto, el nódulo sinusal es prácticamente siempre el marcapasos del corazón normal. Un marcapasos que está situado en una localización distinta al nódulo sinusal se denomina marcapasos ectópico. Un marcapasos ectópico da lugar a una secuencia anormal de contracción de las diferentes partes del corazón y puede producir una debilidad significativa del bombeo cardíaco. Esto se produce porque a veces este fenómeno se presenta en el nódulo AV o en las fibras de Purkinje cuando una de estas estructuras se altera. En ambos casos el marcapasos del corazón se desplaza desde el nódulo sinusal hasta el nódulo AV o las fibras de Purkinje excitadas.
Un marcapasos que está situado en una localización distinta al nódulo sinusal se denomina marcapasos ectópico. Un marcapasos ectópico da lugar a una secuencia anormal de contracción de las diferentes partes del corazón y puede producir una debilidad significativa del bombeo cardíaco. Esto se produce porque a veces este fenómeno se presenta en el nódulo AV o en las fibras de Purkinje cuando una de estas estructuras se altera. En ambos casos el marcapasos del corazón se desplaza desde el nódulo sinusal hasta el nódulo AV o las fibras de Purkinje excitadas.

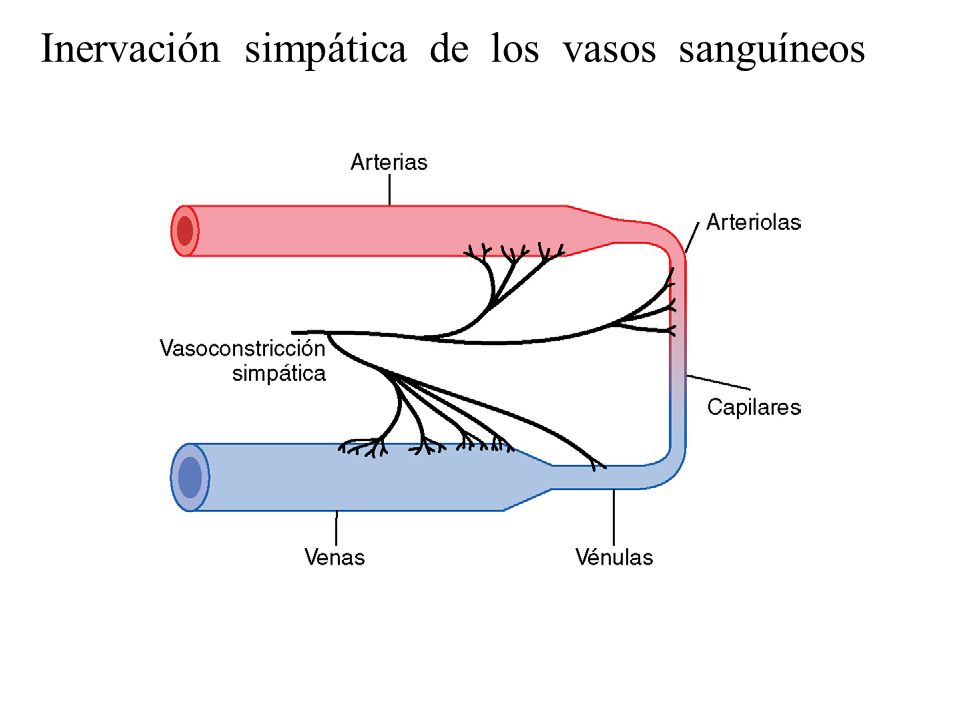
 Los nervios parasimpáticos (vagos) se distribuyen principalmente a los nódulos SA y AV; los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas.
Los nervios parasimpáticos (vagos) se distribuyen principalmente a los nódulos SA y AV; los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas.